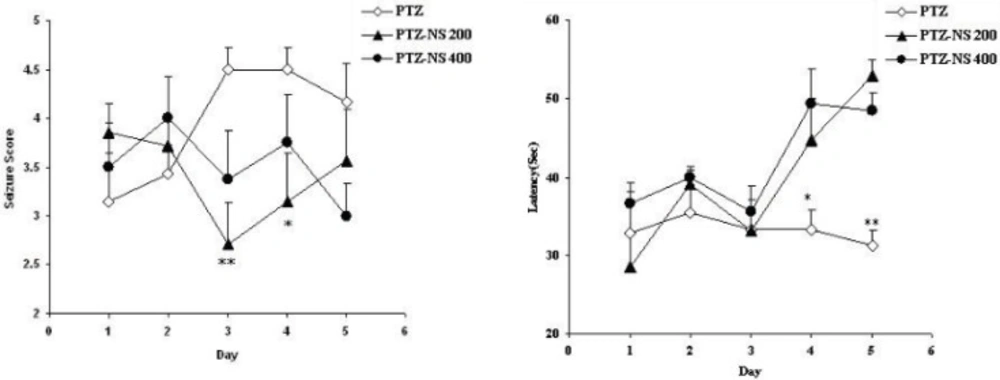
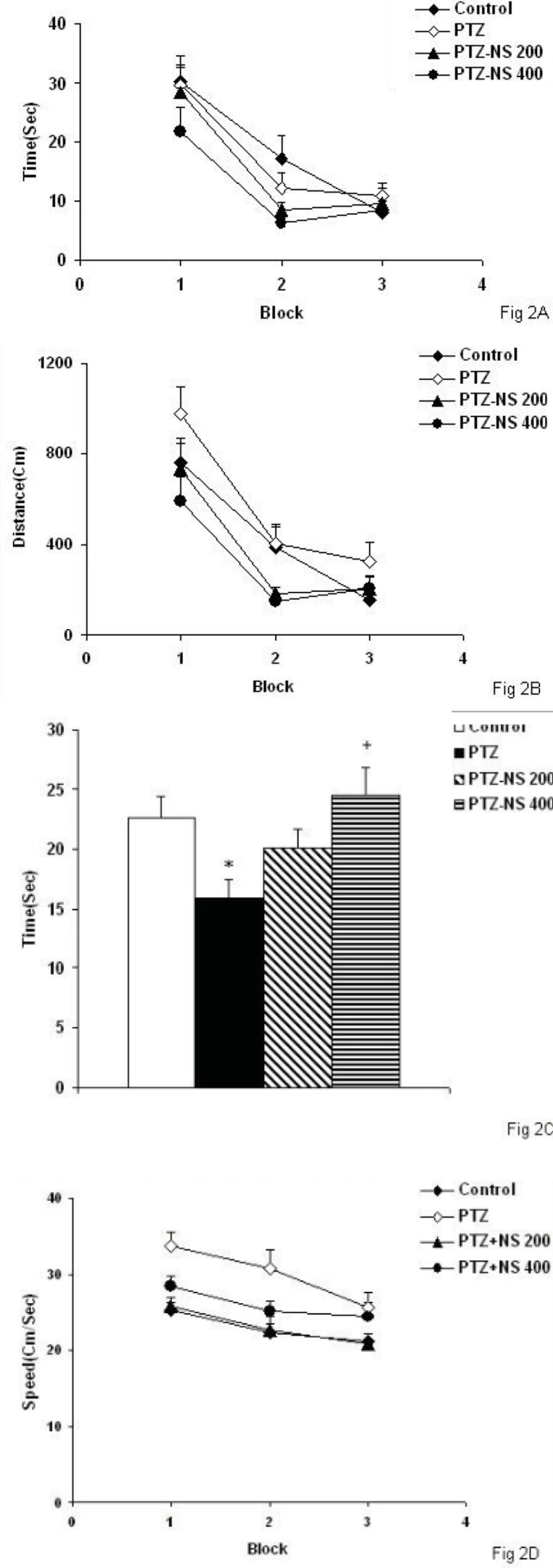
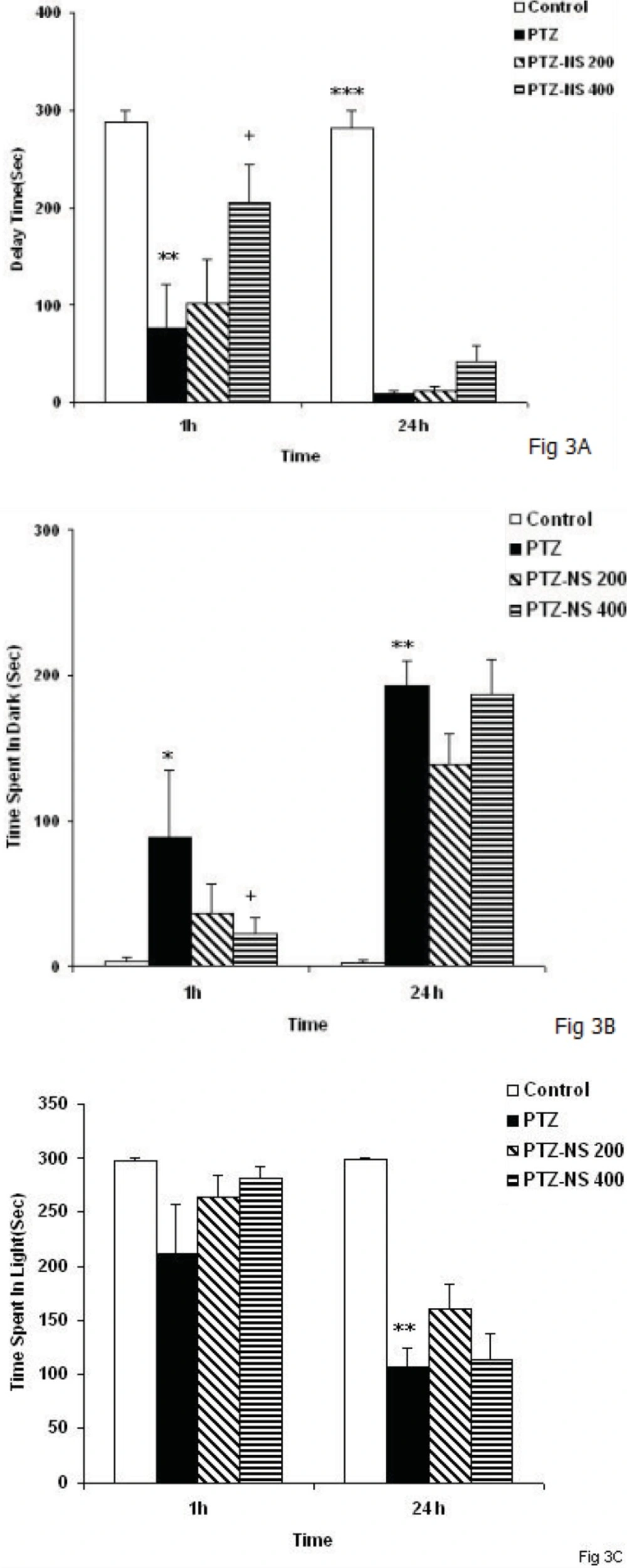
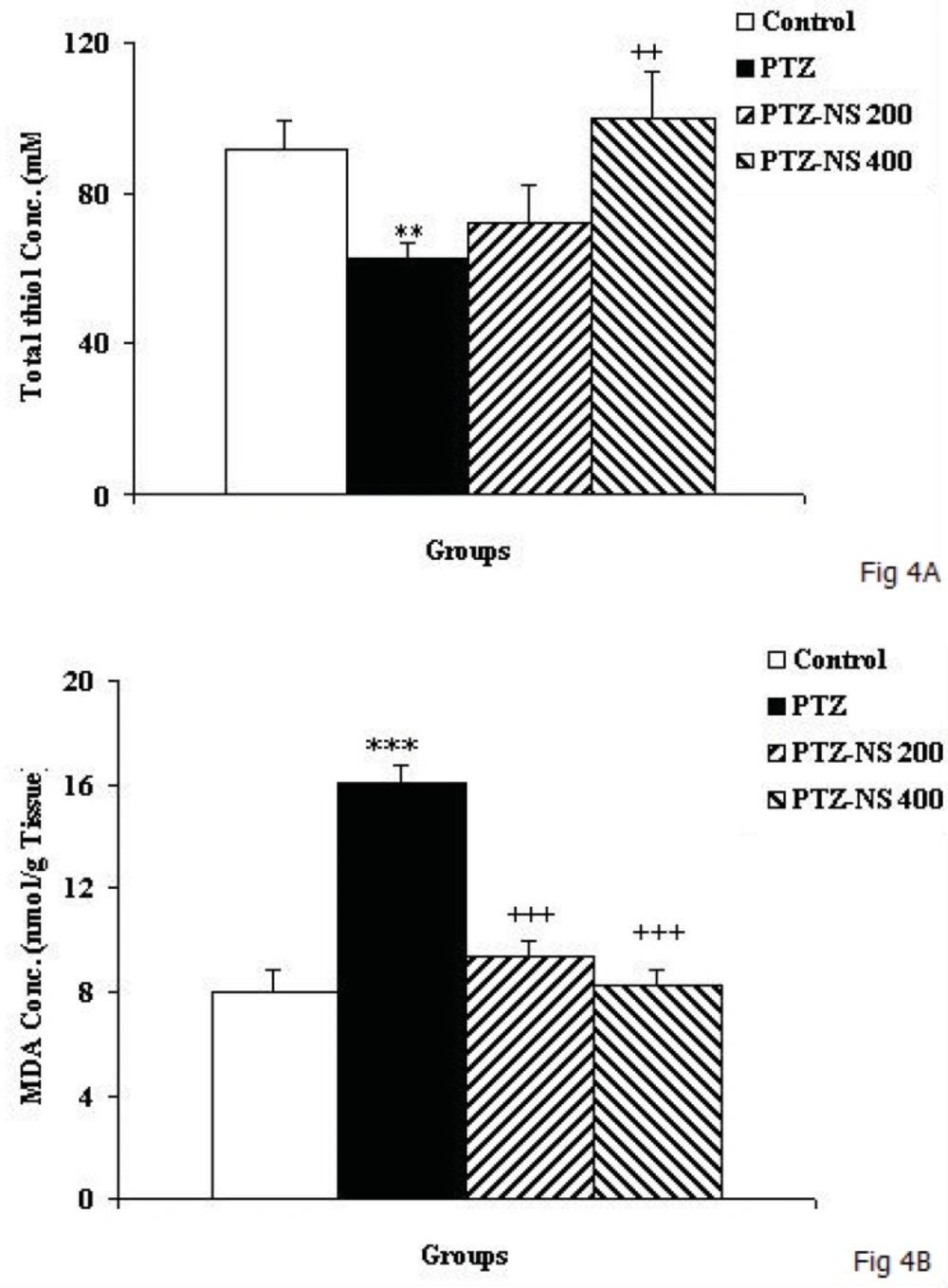
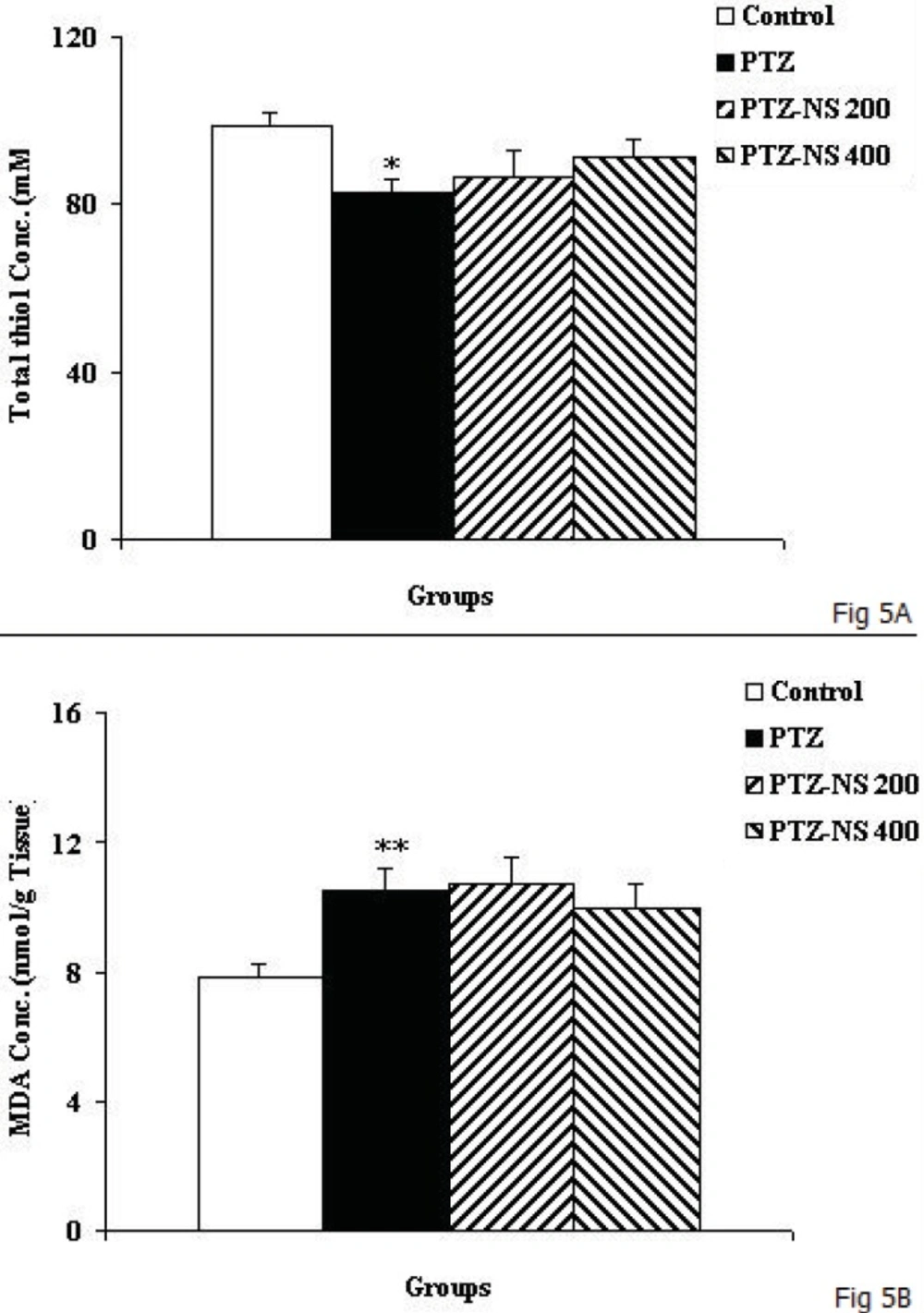
The results of the present study showed that PTZ- induced repeated seizures impaired learning and memory and also induced brain tissue oxidative damage in rats which was reduced by the NS extract. Oxidative stress is a basis for many neurological and neurodegenerative disorders. Oxidative stress is proposed to have a role in the pathogenesis of epilepsy (
1). The presence of a high level of reactive oxygen species (ROS), including superoxide anions, hydroxyl radicals and hydrogen peroxide in the brain after seizures may confirm this idea(
1). However, it has been also well documented that brain tissue oxidative damage has a role in the pathogenesis of the consequences of epilepsy (
2). Accordingly, in the present study an increase in MDA level was observed which was accompanied by a reduction in total thiol groups in the brain tissues of animals subjected to PTZ-induced seizure. The elevation of ROS in the brain may lead to brain tissue oxidative damage which consequently causes problems such as depression, anxiety and memory loss (
32). The results of the present study confirmed that elevation of MDA and decreasing of thiol group concentrations in brain tissues of PTZ treated rats were accompanied with learning and memory impairments. Some study results supported learning and memory decline in epileptic persons (
5). Using kindled animals, it has been repeatedly reported that prolonged and recurrent seizure impairs learning and memory (
33). The results of the present study showed that repeated seizures induced by injection of PTZ for 5 consecutive days also lead to memory impairments. It is also suggested that the effect of repeated seizure isn’t due to its effect on motor activity because its effect on swimming speed was positive in rats but it had a negative effects on memory criteria when the animals were examined in prob test to remember the location of the platform. Regarding the results of the present study the brain tissue oxidative stress as a link between seizures and memory impairments might be suggested.
The seed extract of NS has been used as a natural remedy for treating of a number of illnesses and conditions for many years (
9). The effects of NS on learning and memory have also been suggested. NS oil enhanced learning and memory abilities of the rats when examined using eight-arm radial arm maze (
14). Using Morris water maze also, NS improved learning and memory impairments due to global cerebrovascular hypoperfusion in rats (
12). Protective effects of Nigella sativa against diabetes induced learning and memory impairment has also been reported (
15). Results from human studies also confirmed the beneficial effects of NS on memory, attention and cognition without any side effects on kidney, heart and liver (
13)
Mohamed
et al. (2005) showed that treatment of rats with thymoquinone prevented the experimental autoimmune encephalomyelitis (
34), which was probably due to its antioxidant and anti-inflammatory properties (
35). The antioxidant effects of NS and thymoquinone have been repeatedly reported to be comparable to the effects of vitamine E and vitamine C (
23,
25). However it was not compared to other well-known antioxidants, the results of the present study also showed that pretreatment by NS extract prevented elevation of MDA brain tissue concentration while it enhanced total thiol concentration.
Moreover, it has been previously reported that hippocampal neurodegeneration after chronic toluene exposure in rats was prevented by
NS oil and thymoquinone (
36). Hosseinzadeh
et al. have also shown that NS and thymoquinone prevented lipid peroxidation increment in hippocampal proteins following global cerebral ischemia-reperfusion injury model in rats (
25). The anti-Parkinson’s activity of NS was also accompanied with the antioxidant effects (
27,
37). The antioxidant effects of NS oil was comparable to the effects of valproate in PTZ induced kindled mice (
26). Antiepileptic effect of NS and its ability to inhibit excessive ROS formation in PTZ - induced seizures has been reported (
26). A neuroprotective effect of NS in an experimental model of spinal cord injury in rats has also been attributed to its antioxidant capability (
38). It has been shown that NS extract and thymoquinone attenuated oxidative stress and neuropathy in diabetic rats (
39). Regarding these reports and the results of present study, it seems that NS attenuates both seizures severity and brain tissues oxidative damage and protects learning and memory impairments in rats. It does not seem that the effect of the extract be due to its effect on motor activity because both doses of the extract reduced the speed compared to PTZ group but they enhanced the time spent in target quadrant in prob test.
NS was also found that modulates the brain content of excitatory and inhibitory neurotransmitters such as aspartate and glutamate, GABA and glycine (
40). NS increased GABA released from cultured neurons whereas secretion of glutamate, aspartate, and glycine were decreased. (
41). Regarding these findings and the sedative and depressive effects of NS observed
in-vivo, the changes of inhibitory/excitatory amino acids levels may have a role in the effects which was seen in the present study. The effects of NS and thymoquinone on pentobarbital-induced hypnosis, locomotor activity and motor coordination which have been previously reported might also be conceivable (
18).
Hosseinzadeh and Parvardeh using a PTZ model showed that thymoquinone prolonged the onset of seizures and reduced the duration of myoclonic seizures and reduced mortality rate however, in maximal electroshock (MES) model, thymoquinone failed to reduce the duration of seizure, whereas exhibited a complete protection against mortality. In this study the interactions with opioid and GABAergic systems were suggested as possible mechanisms (
18). However, thymoquinone didn’t show the protective activity on percentage of protection of animals against MES- induced tonic seizures in another study (
42). Thymoquinone has been reported to have anti-epileptic effects in children with refractory seizures (
19). In another study NS aqueous extract, and its volatile oil and hymoquinone, showed antiepileptic effects using PTZ induced seizures model and potentiated the effects of valproate (
43). Nigella sativa oil reduced the sensitivity of kindled mice to the convulsive effects of PTZ and mortality rate while, valproate was ineffective in preventing development of any of these effects (
26). It was also shown that thymoquinone in both PTZ and MES- induced seizure models increased sodium valproate potency and also reduced the hepatotoxic effects of sodium valproate (
44)
.Furthermore, it has been postulated that
Nigella sativ
a oil and thymoquinone produced antinociception in the formalin test by releasing endogenous opioid peptides in the CNS, and causes an antinociceptive tolerance following repeated administration (
45). It was also shown that the antinociceptive effect of morphine was reduced in thymoquinone and NS oil-pretreated mice (
45). The ability of the hydroalcoholic extract of NS to change morphine induced conditioning place preference may also confirm the interaction of the plant with opioid system (
46). Therefore, it might be suggested that NS and its ingredients have interaction with opioid system to reduce seizure severity and prevent memory impairments which was seen in the present study but it needs to be more investigated.
As it was mentioned NS is containing many compounds, the main of them is thymoquinone. In the present study the ingredient(s) responsible for the effects of the extract was not known and it needs to be investigated in future. Recently, it is reported that besides thymoquinone, the main ingredient which has an important role in pharmacological effects of NS, other component(s) may also be involved in the results of the present study. More recently, melanin has been shown to occur abundantly in the seed coats of NS (
47). On the other hand, according to a number of botanical sources, melanin has been found to act as an immunomodulator (
48,
49). Therefore, involvement of this ingredient of NS may also be suggested as a possible compound which plays a role in the result of the present study.