Osteosarcoma is a primary bone malignancy with high rates of metastasis, mortality and disability, and this disease usually occurs in children and adolescents (
1). The most common treatments for osteosarcoma are surgery, chemotherapy and biotherapy. However, the prognosis of osteosarcoma is still need to be further investigated because of the high degree of malignancy, rapid disease progression and early metastasis (
2). Though the traditional treatments have been improved (such as surgery and chemotherapy), the efficacies of new therapy methods are still needed to be evaluated to improve survival rate. With the developments of natural medicinal chemistry and molecular biology, the fundamental research of osteosarcoma has also been promoted. Traditional Chinese herbal extract is one of the research hotspots nowadays (
3). The therapeutic strategies that combine traditional Chinese medicine or its active ingredients with surgery and chemotherapy treatment can reduce the toxicities of traditional therapy and drugs, leading to the improvements of patients' immunity and compliance (
4-
6).
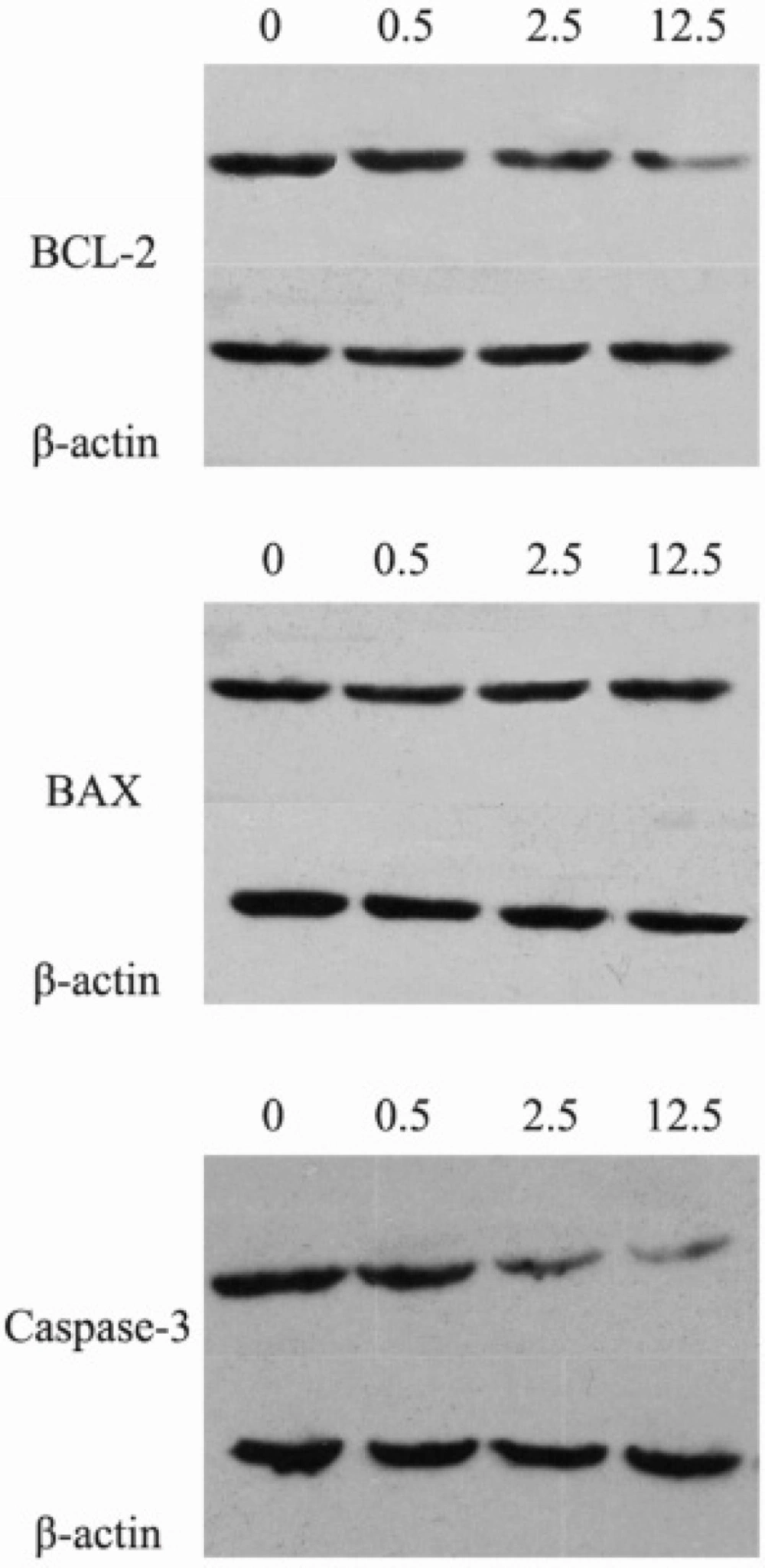
BCL-2 (B-cell lymphoma 2), the founding member of the BCL-2 family, participates in the regulation of cell apoptosis (
7). BAX (Bcl-2-associated X protein), a member of the BCL-2 protein family, promotes cell apoptosis by binding to and antagonizing the BCL-2 protein. The BAX gene is the first identified pro-apoptotic Bcl-2 family member (
8). Damage to the BCL-2 gene has been identified as a cause of many cancers, including melanoma, breast cancer and prostate cancer. The Caspase 3 protein, a member of the cysteine-aspartic acid protease (caspase) family, is a crucial mediator of programmed cell death (
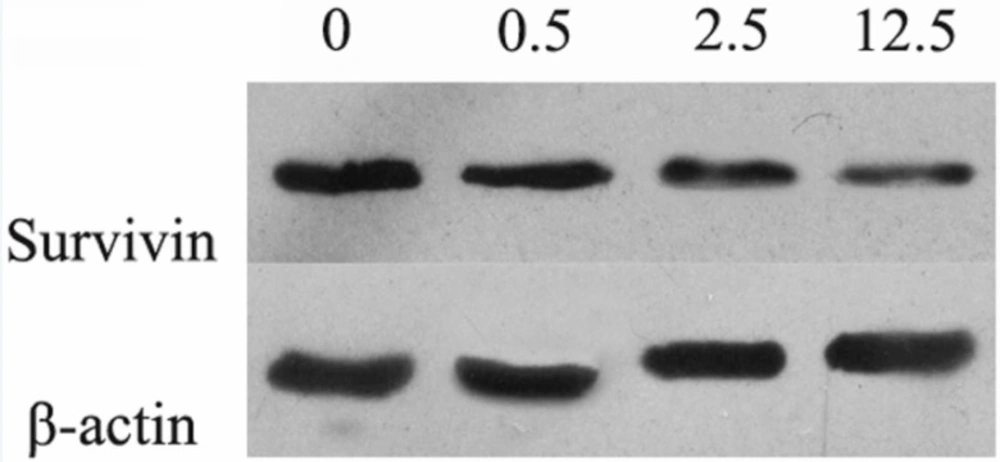
9). Survivin, a member of the inhibitor of apoptosis (IAP) family, could inhibit caspase activation to negative regulate apoptosis or programmed cell death. Luteolin (3',4',5,7-tetrahydroxyflavone) is a natural flavonoid compound extracted from a variety of traditional Chinese medicines, such as
Lonicerajaponica,
Schizonepetatenuifolia and
Ajugadecumbens Thunb (
2). The important structure features of luteolin, including 2-3 carbon double bond and hydroxyl moieties at carbons 5, 7, 3’, and 4’ positions, are associated with the biological and biochemical activities of luteolin (
10,
11). The anticancer property of luteolin is related to the biological effects of luteolin, such as angiogenesis, anti-inflammatory activity, inhibition of cell proliferation, induction of apoptosis and metastasis (
12). Previous reports have indicated that luteolin has various biological activities, such as immunoregulation (
13), anti-oxidant (
14), anti-inflammation (
15), cardiovascular protection (
16) and enzyme inhibition (
17). In recent years, luteolin has been proved to inhibit the proliferation of leukemia cells (
18) and invasion of prostate cancer cells (
19,
20). Luteolin also could induce the apoptosis of hepatoma cells (
21,
22) and lung cancer cells (
23). What’s more, luteolin has anti-proliferative, chemosensitizing and radiosensitization effects on gastric cancer cells (
24,
25). However, the effects of luteolin on human osteosarcoma cells are still unknown.
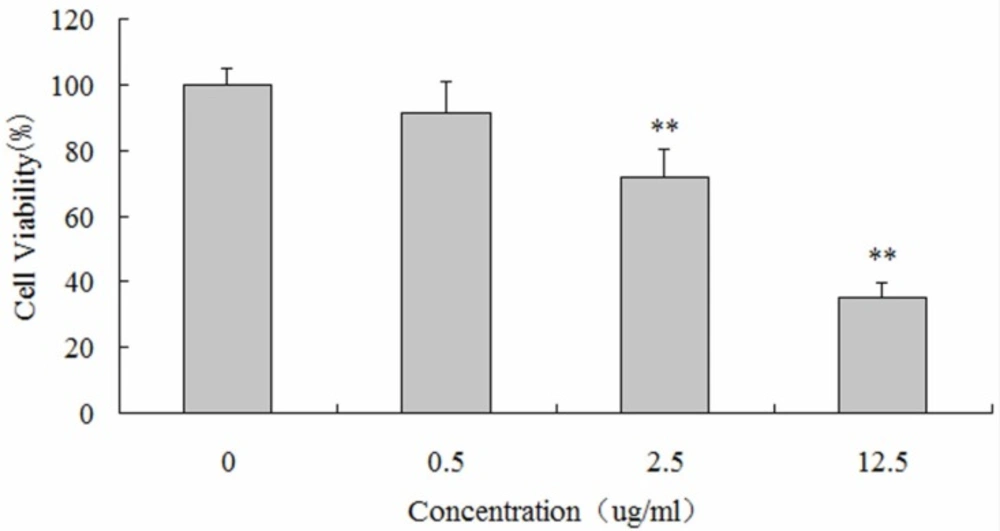
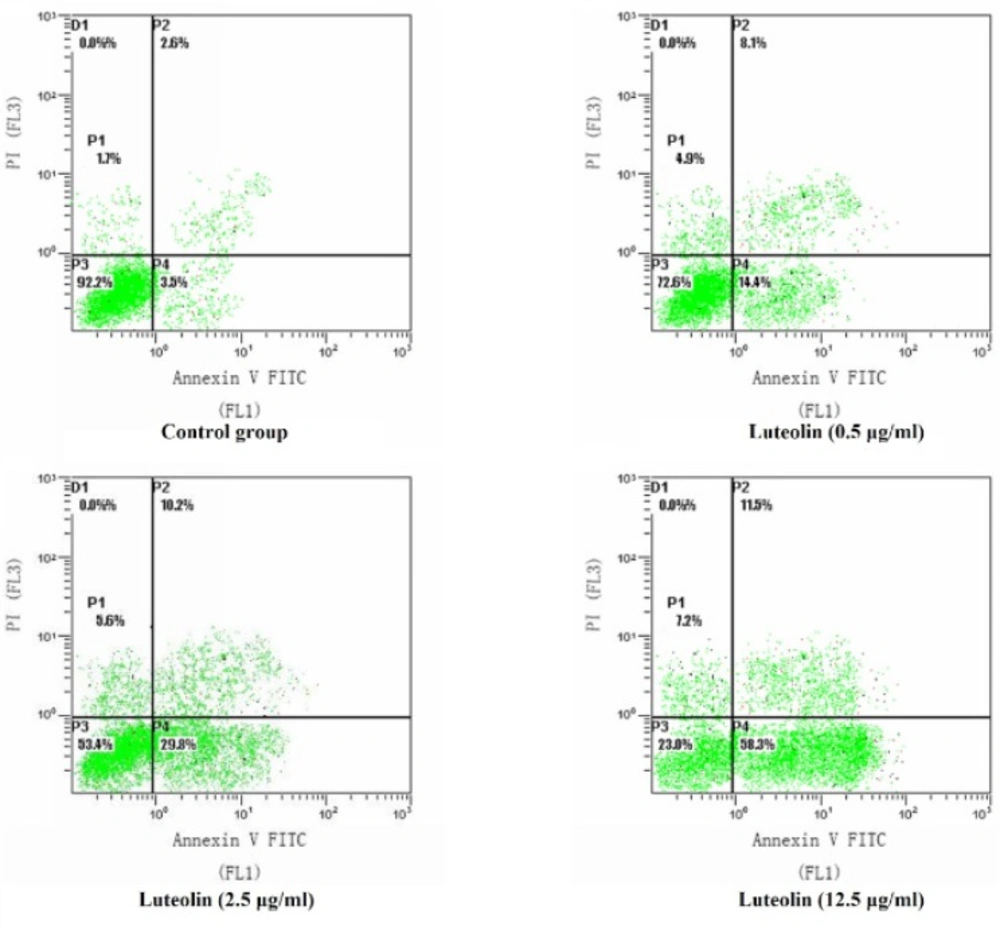
This study was undertaken to investigate the effects and molecular mechanisms of luteolin on the proliferation and apoptosis of osteosarcoma cells. We established osteosarcoma cell model in-vitro and intervened osteosarcoma MG-63 cells with luteolin. Not only the proliferation and apoptosis of MG-63 cells, but also protein expressions of BCL-2, BAX, Caspase-3 and Survivin were measured.