Animals
Male Wistar rats (Pasteur Institute; Tehran, Iran) weighing 200-230 g at the time of training were used. The animals were kept in an animal house with a 12-h light/ 12-h dark cycle and controlled temperature (22 ± 2°C). Availability of food and water to animals was ad libitum. Animals were kept in an animal room for at least 1 week to adapt to the laboratory conditions before training. All procedures were performed in accordance with institutional guidelines for animal care and use. The Research and Ethics Committee of the college of science at the University of Tehran approved the experimental protocol.
Passive avoidance apparatus
Passive avoidance apparatus for training and testing contained a box of two compartments, one light (white compartment) and the other dark (dark compartment) with the same size (20 × 20 × 30 cm)that were connected to each other with a guillotine door (7 × 9 cm). The floor of the dark compartment was replaced with stainless steel rods (2.5 mm in diameter) at 1cm intervals to produce foot shock. Intermittent electric shocks (50 Hz, 3s, 1mA) were delivered by an insulated stimulator (Borj Sanat, Iran).
Training and testing
Training
Training was performed 30 min after habituation of the animals in the experimental room. Each animal was gently placed in the brightly lit compartment of the apparatus; the guillotine door was then opened after 5 s to let the animal enter the dark compartment. The latency with which the animal crossed into the dark compartment was recorded. Any animal waiting more than 100s in the light compartment was eliminated from the study. Once the animal entered the next compartment with all four paws, the door was closed and the animal was taken to its home cage. The trial was repeated after 30 min (acquisition trial), in which a foot shock (50 Hz, 1 mA, 3 s) was immediately delivered to the grid floor of the dark room. The rat was removed from the apparatus and placed temporarily into its home cage. Two minutes later, the procedure was repeated; if the rat did not enter the dark compartment during 120 s, it was recorded as successful acquisition of passive avoidance response. Otherwise, when the rat entered the dark compartment before 120 s, the door was closed and the same shock was applied again. Upon retesting, if the rat acquired passive avoidance successfully, it was removed from the apparatus and put back to its home cage.
Testing
Retrieval test was performed 24 hours after training to determine long-term memory. Each animal was gently placed in the light compartment; the door was opened and the step through latency was measured for entering into the dark compartment. Testing session ended when the animal entered the dark compartment or remained in the light compartment for 300 s (criterion for retrieval). During testing session, no electric shock was applied.
Drugs
The drug used in the study was morphine sulfate (Temad Co.,Teharan, Iran). Morphine was dissolved in sterile 0.9% saline. Immediately after successful training, morphine was injected subcutaneously (post-training s.c. injection). Control animals received saline subcutaneously (s.c.).
Experimental procedure
Effect of morphine on memory retrieva lin passive avoidance task
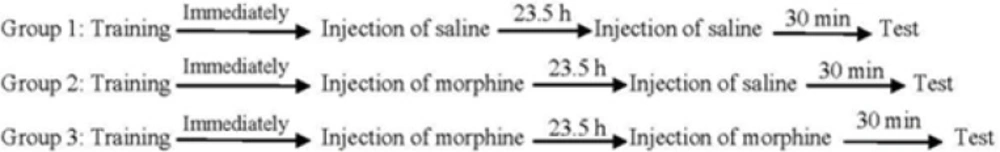
Three experimental groups of eight animals were used in order to evaluate the effects of post-training and/or pre-test administration of morphine on memory retrieval of passive avoidance task. One group of animals received saline (1 mL/Kg,
s.c.) immediately after training (post-training). On the test day, the animals received saline (1 mL/Kg,
s.c.) 30 min before the test (pre-test). Another two groups of animals received post-training injection of morphine (7.5 mg/Kg,
s.c.) and after 23.5 h, they received pre-test injections of saline (1 mL/Kg,
s.c.) or morphine (7.5 mg/Kg,
s.c.). The step-through latency was measured 30 min after the injection (
Figure 1).
Summery of experimental design in passive avoidance task.
Hippocampus extraction
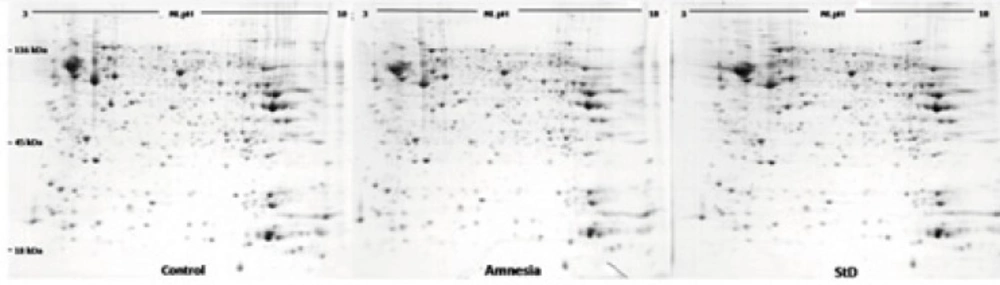
In order to perform hippocampal proteomic analysis, three animals with the best step-through latency score were randomly chosen from each group. Each animal was sacrificed by rapid guillotine decapitation after the test session. The brain was removed from the skull and the hippocampus extracted quickly and easily from rats as described by Chiu and co-workers (2007) (34). The hippocampus was frozen in liquid nitrogen before storing at -80 ˚C.
Protein extraction
Protein extraction was performed as described by Hirano
et al. (
35) with some modifications. Trichloroacetic acid (TCA)/acetone extraction buffer (AEB) was used for protein extraction. TCA-AEB is a suitable solution for protein extraction which denatures proteins and inactivates enzymes (see
Table 1). AEB is used to eliminate the interference effect of TCA in isoelectric focusing (IEF). It should be considered that one of the most important considerations in two-dimensional gel electrophoresis (2DE) is the preparation of
protein samples. All the buffers and detergents should be compatible with IEF. The choice of material for protein extraction requires considerations of salt concentration and protein solubility, because high concentration of salt and protein precipitation can be problematic. Among protocols, TCA-AEB is the most suitable extraction method for reliable IEF. Each hippocampus was homogenized separately with pestle in TCA/AEB, and centrifuged at 15000 rpm, 15 min, at 4 ˚C after incubation for 1 h at -20 ˚C. Lysis buffer (7M Urea, 2M thiourea, 4% CHAPS, 18 mM Tris-HCL (PH 8.0), 50 mM DTT) was added to the pellet and centrifuged at 15000 rpm for 20 min at 10 ˚C. The protein concentration of the supernatant was estimated (20-30 µg/µL) through Bradford method (
36) using NanoDrop 2000c Thermo Spectrophotometer.
| Material | Amount | Protein extraction solution |
|---|
| tricholoroacetic acid (TCA) | 10% | TCA-AEB |
| Merchaptoethanol | 0.07% |
| Acetone | To needed volume |
| Merchaptoethanol | 0.07% | AEB |
| Acetone | To needed volume |
| Urea | 7M | Lysis buffer |
| Thiourea | 2M |
| CHAPS | 4% (v/v) |
| Tris-HCL (PH 8.0) | 18 mM |
| Dithioteriol (DTT) | 50 mM |
| Ultra pure water | To needed volume |
Two-dimensional gel electrophoresis
The first dimension started with Rehydration step in which a Rehydration buffer (8 M urea, 4 % CHAPS, 0.2 % ampholyte pH 3-10, 50 mM DTT ) containing 1mg protein sample was applied onto Non-linear immobilized pH gradient strips (NL IPG, pH 3-10, 17cm). Isoelectric focusing (IEF) was carried out with Protean IEF Cell (BioRad, USA) at 50000 Vh. Before starting the second dimension, Equilibration buffer (6 M Urea, 2% SDS, 50 mM Tris-HCL (pH 8.8), 20% v/v Glycerol) containing 2% DTT (first step) or 2.5% Iodoacetamide (second step) was applied onto focused strips for 20 min. The second dimension was carried out according to Laemmli buffer system (
37), using handmade 12% SDS-polyacrylamide gels and run in Protean II xi Cell (bioRad, USA). Each gel was placed separately in bluesilver staining solution for one night (
38).
Image analysis
Analytical gels were scanned by a Densitometer GS-800 (BioRad) scanner at 300 dpi in tagged image file format (TIFF). Image Master™ 2D Platinum v6.0 software was then used to extract and digitize data from graphical images of scanned gels through detecting, normalizing, matching and comparing protein spots according to their volume percent. Student t-test was performed and spots with more than 1.5 fold changes in vol% selected for identification by mass spectrometry and statistical significance was p<0.05.
In-gel trypsin digestion of proteins
Spots were manually cut from 2-DE gels, placed in 96 wells v-shape polypropylene plates and dried completely. The Ettan Spot Handling Workstation (GE Healthcare, UK) was used for automatic in-gel digestion of samples. Each gel plug was soaked in 100 µL of washing solution (50% MeOH, 50 mM NH4HCO3) to re-swell and was then washed two more times in the same solution. The gel plugs were further washed twice in 75% ACN, before being completely dried. Samples were then re-hydrated by adding freshly prepared trypsin solution (0.5 µg modified porcine trypsin in 25 µL 20 mM NH4HCO3), and were incubated for 240 min at 37 oC. Peptides were extracted from the gel plugs, by washing twice in 100 µL of 50% ACN, 0.1% TFA and transferred in solution to a fresh 96 well plate, where samples were dried.
Matrix-assisted laser desorption ionization time-of-flight mass spectrometry (MALDI-TOF/TOF-MS)
Tryptic peptides were resuspended in 3 µL of 50% ACN, 0.1%TFA. 0.3 µL of resuspended tryptic peptides were spotted onto a steel Applied Biosystems 192 sample MALDI target plate, and were mixed (while wet) with 0.3 µL of a 90% saturated µ-cyano-4-hydroxycinnamic acid (CHCA) in 50% ACN, 0.1% TFA. The dried samples were analyzed using a MALDI-TOF/TOF MS (4700 Proteomics Analyzers, Applied Biosystems, UK), performing MS analysis and subsequent MS/MS analysis on up to 10 precursor peptides. Each sample was internally calibrated by reference to specific autolytic fragements of trypsin. Mass tolerance settings of 1.2 Da for parent ion and 0.5 Da for fragment ions were applied. Search settings allowed one missed cleavage with trypsin and two modifications (carboxamidomethylation of cysteine and oxidation of methionine). The PMF and MS/MS information were automatically searched against the NCBI non-redundant database using the Mascot search engine (Matrix Science, UK). The Mascot score cut-off value was set to 88 for the statistical confidence limits of 95%.
Statistical analysis of memory retrieval
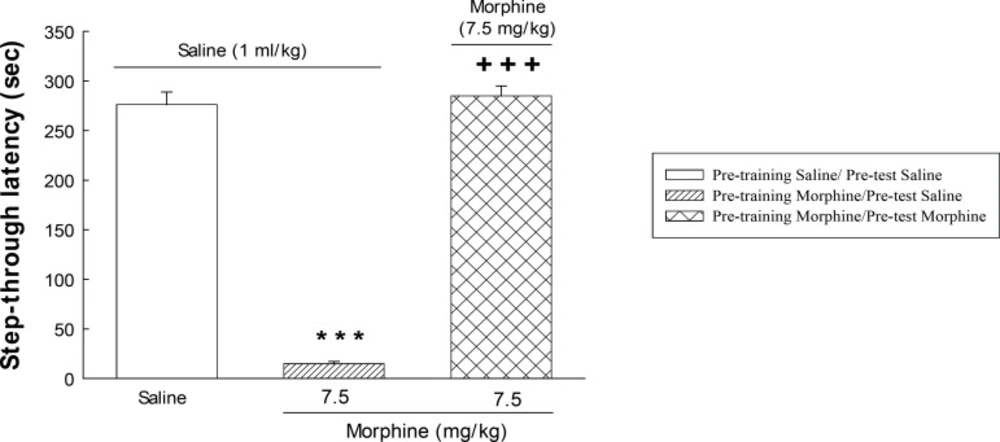
The step-trough latencies as an index of memory retrieval (
Figure 2) are expressed as means ± S.E.M. The statistical analysis was performed using one-way analysis of variance (ANOVA). The level of statistical significance was set at p < 0.05. Post-hoc comparison of means was carried out using Tukey test for comparisons. Calculations were performed using the SPSS statistical package(Version 20).
The effect of post-training administration of morphine on step-through latencies. The animals received post-training saline (1 mL/Kg, s.c.) or morphine (7.5 mg/Kg) and were tested after 24 h. Each value represents the mean ± S.E.M. of eight rats per group. ***P < 0.001 compared with the saline/saline group.+++P < 0.001 compared with the morphine/saline group.