Adult male Wistar rats (190-250 g; n = 50) (Pasteur’s Institute, Tehran, Iran) were housed three to four per cage in a temperature-controlled colony room under light/dark cycle with food and water available ad libitum. Procedures involving animals and their care were conducted in conformity with NIH guidelines for the care and use of laboratory animals. The animals were held in the colony room for at least two weeks before being tested. Only rats not showing any biased rotational behavior (net rotations less than 30/hour) following intraperitoneal injection of apomorphine hydrochloride (2 mg/Kg) (Sigma Chemical, USA) were selected for the present study. The animals were randomly divided into five groups: sham-operated group, thymoquinone10-treated sham-operated groups (Sham + Thymoquinone10), lesion group (6-OHDA) and thymoquinone-treated lesion groups (6-OHDA + Thymoquinone5 and 6-OHDA + Thymoquinone10). Unilateral intrastriatal 6-OHDA (Sigma Chemical, USA) injection (left side) was performed through a 5 μL Hamilton syringe on anesthetized rats (ketamine 80 mg/Kg and xylazine 10 mg/Kg,
i.p.) using stereotaxic apparatus (Stoelting, USA) at the coordinates: L –3 mm, AP 9.2 mm, V 4.5 mm from the center of the interaural line, according to the atlas of Paxinos and Watson (
14). At the end of injection, the needle was left in place for an additional 5 min and then withdrawn at a rate of 1 mm/min. The lesion group received a single injection of 5 μL of 0.9% saline containing 2.5 μg/μL of 6-hydroxydopamine-HCL (6-OHDA, Sigma Chemical, USA) and 0.2% ascorbic acid (W/V) at a rate of 1 μL/min. The sham group received an identical volume of ascorbate-saline solution. The 6-OHDA + thymoquinone5 and 6-OHDA + thymoquinone10 groups received the neurotoxin in addition to TQ p.o. (using rodent gavage) dissolved in propylene glycol (Merck, Germany) at doses of 5 and/or 10 mg/Kg respectively. TQ (Sigma Chemical, USA; purity > 97%) was daily administered from two days before surgery with an interval of 24 h (
3). The third injection of thymoquinone was 1 h before surgery.
Behavioral testing
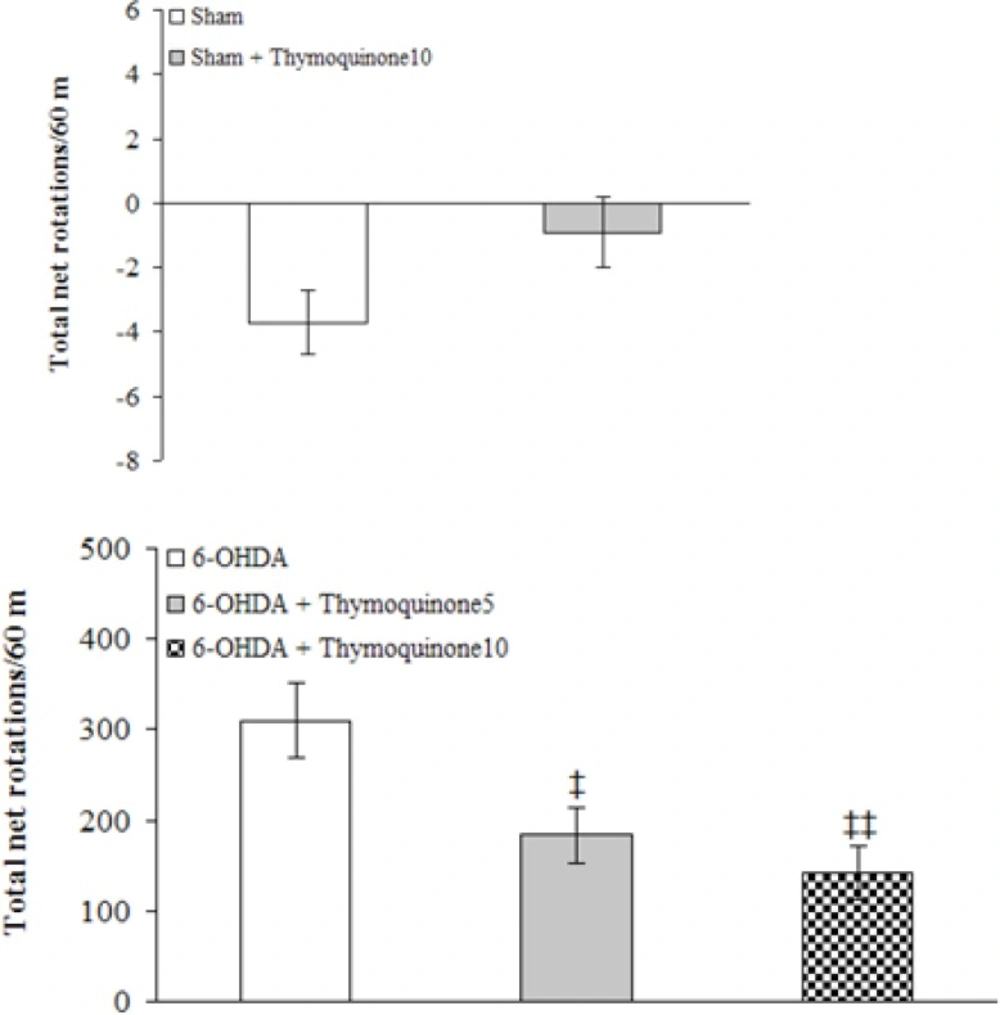
The animals were tested for rotational behavior by apomorphine hydrochloride (2 mg/Kg,
i.p.) one week before surgery (baseline) and after 1 week. The rotations were measured according to a method as described previously (
3). Briefly, the animals were allowed to habituate for 10 min and then 1 min after the injection, full rotations were counted in a cylindrical container (a diameter of 33 cm and a height of 35 cm) at 10-min intervals for 60 min in a dimly-lighted room. Net number of rotations was defined as the positive scores minus the negative scores.
Determination of midbrain MDA concentration
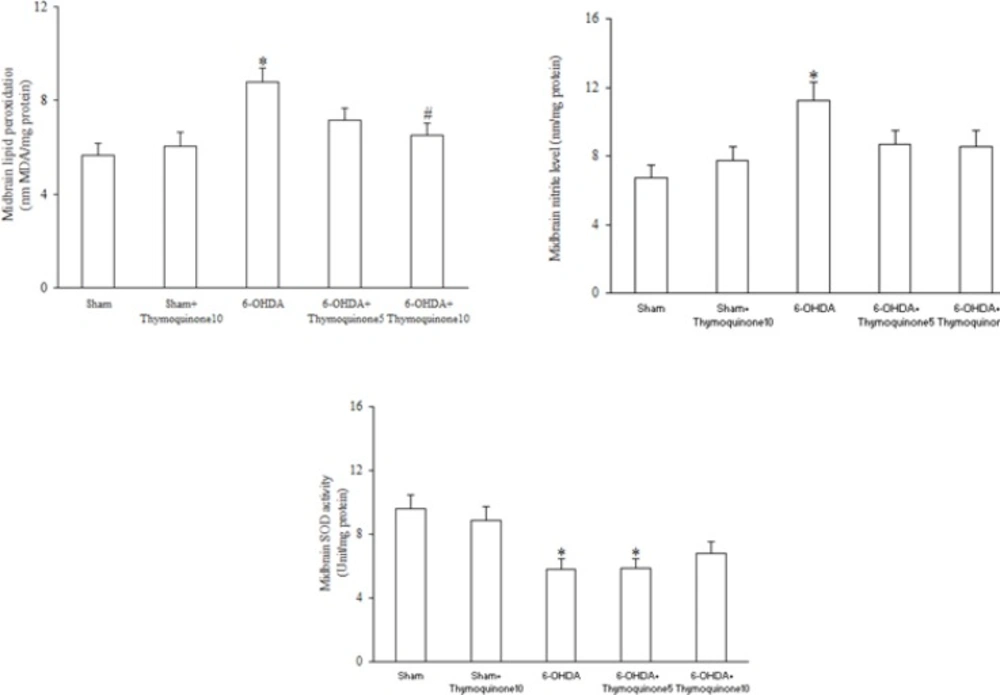
The rats were anesthetized with ketamine (150 mg/Kg), decapitated, brains were removed, anterior third block of left midbrain was blotted dry, weighed, then made into 5% tissue homogenate in ice-cold 0.9% saline solution, centrifuged at 4 ºC, obtained supernatant was aliquotted, then stored at −80 °C until assayed (
3). The MDA concentration (thiobarbituric acid reactive substances, TBARS) in the supernatant was measured as described before (
3). Briefly, trichloroacetic acid and TBARS reagent were added to supernatant, then mixed and incubated at boiling water for 80 min. After cooling on ice, samples were centrifuged at 1000×g for 10 min and the absorbance of the supernatant was read at 532 nm. TBARS results were expressed as MDA equivalents using tetraethoxypropane as standard.
Measurement of midbrain SOD activity
The supernatant of midbrain homogenate was obtained as described earlier. SOD activity measurement was according to previous works (
3). Briefly, supernatant was incubated with xanthine and xanthine oxidase in potassium phosphate buffer (pH 7.8, 37 ºC) for 40 min and NBT was added. Blue formazan was then monitored spectrophotometrically at 550 nm. The amount of protein that inhibited NBT reduction to 50% maximum was defined as 1 nitrite unit (NU) of SOD activity.
Assay of midbrain nitrite concentration
Supernatant nitrite content was assayed by the Griess method according to previous studies (
15). Because NO is a compound with a short half-life and is rapidly converted to the stable end products nitrate (NO3 -) and nitrite (NO2–), the principle of the assay is the conversion of nitrate into nitrite by cadmium and followed by color development with Griess reagent (containing sulfanilamide and N-naphthyl ethylenediamine) in acidic medium. The total nitrite was measured by Griess reaction. The absorbance was determined at 540 nm with a spectrophotometer.
Protein Assay
The protein content of the supernatant was measured with Bradford method using bovine serum albumin (Sigma Chemical, USA) as the standard (
16).
Histological study
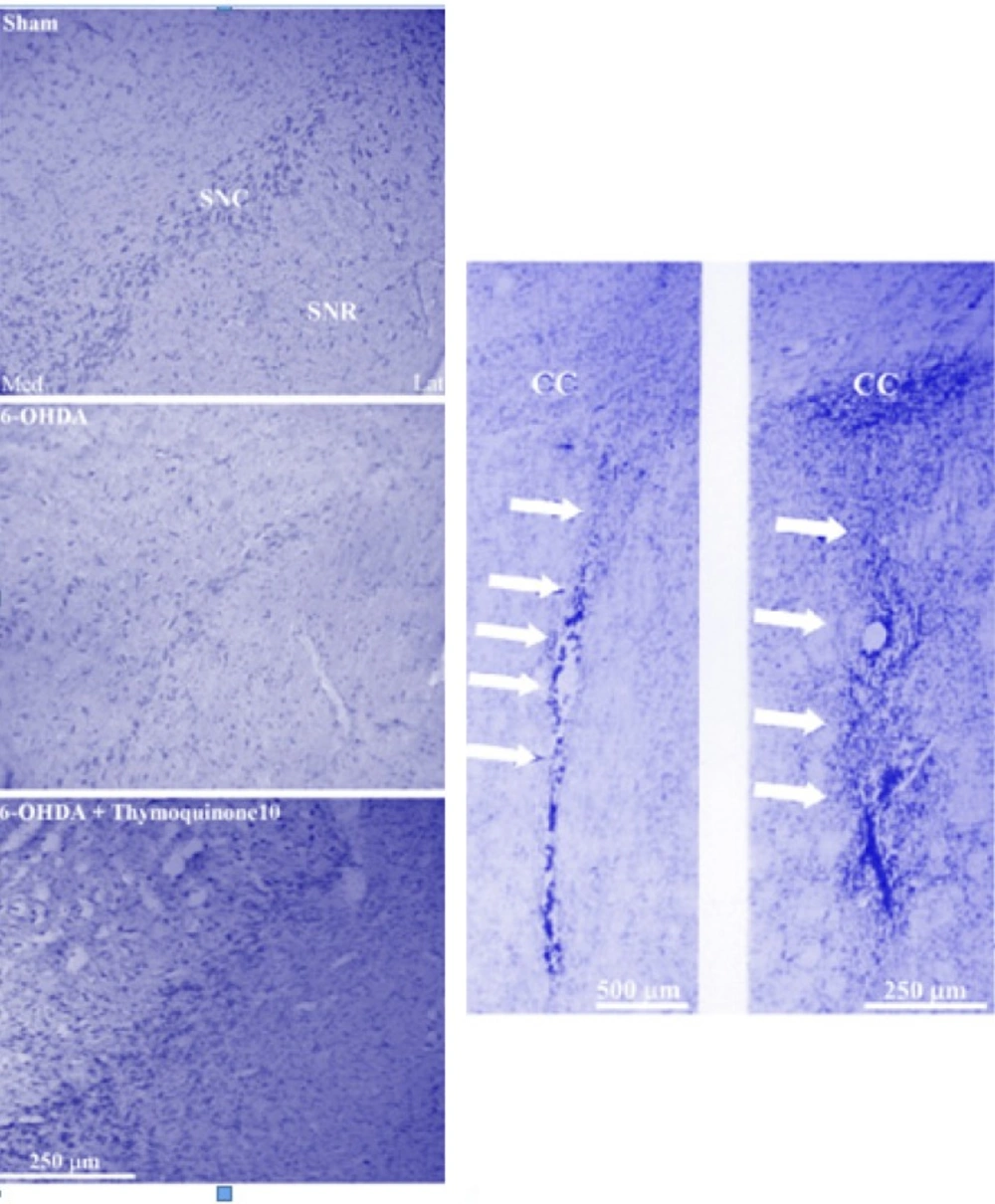
Half of the animals in each group were randomly used for histological assessment. At the end of behavioral experiments, the rats were deeply anesthetized with a high dose of ketamine (150 mg/Kg) and perfused through the ascending aorta with 50-100 mL of 0.9% saline followed by 100-200 mL of fixative solution containing 4% paraformaldehyde in 0.1 M phosphate buffer (PB, pH 7.4) followed by 100 mL of 0.1 M PB containing 10% sucrose. Following perfusion, the brains were removed from the skull, blocks of forebrain and brainstem were prepared, and after final steps of preparation (immersion in 30% sucrose solution for 2-3 days), sections were cut at a thickness of 40 μm on a freezing microtome (Leica, Germany) and collected in PB (0.1 M). Every second section was Nissl-stained with 0.1% cresyl violet (Sigma chemical, USA).
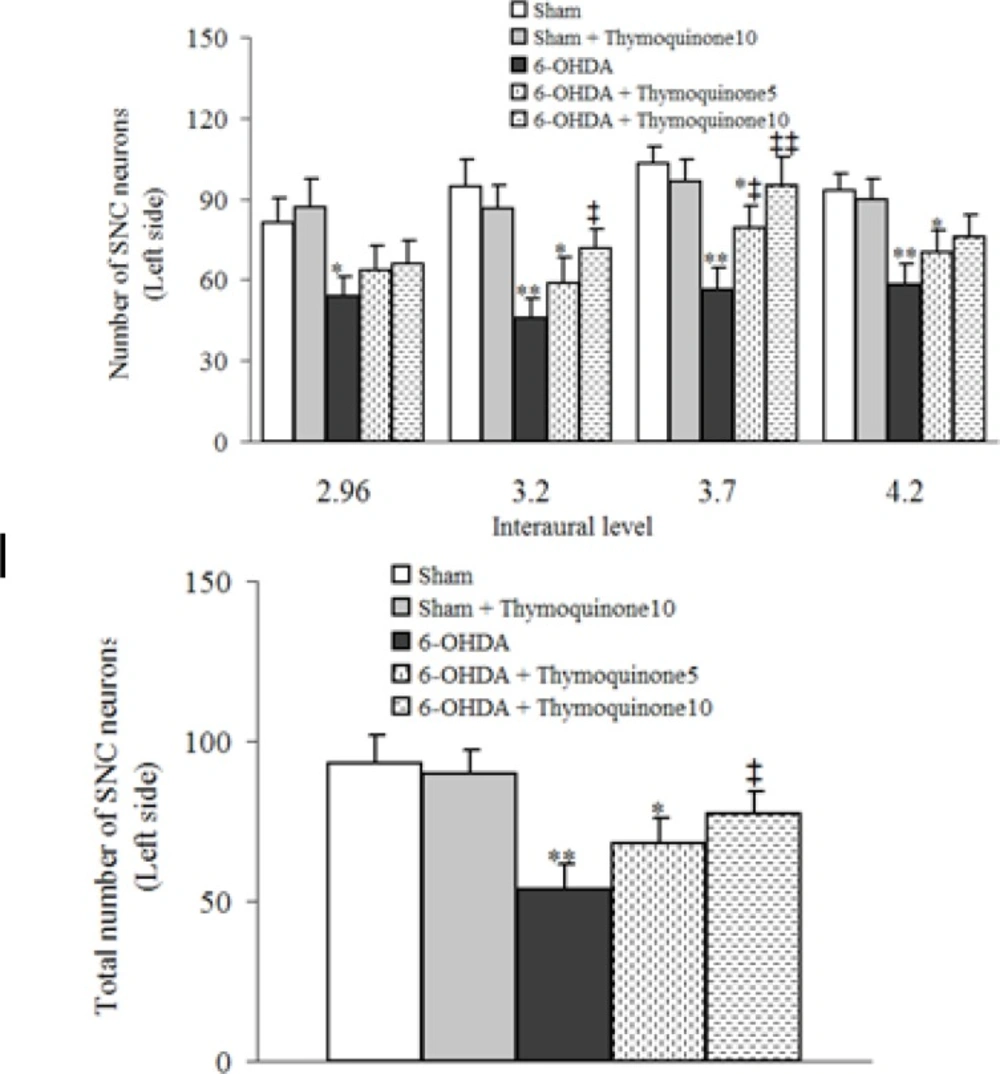
Neuronal counting
For each animal, mesencephalic sections (Interaural 2.9-4.2 mm) were examined by a method as described previously (
17). Briefly, Nissl-stained neurons of the SNC were counted manually (Light microscopy; X400) using a superimposed grid to facilitate the procedure. At least two sections representative of each of four Paxinos-Watson planes (4.2, 3.7, 3.2, 2.97; Interaural) were examined by scanning the entire extent on each side. Counting was done blind to the treatments received.
Statistical analysis
All data were expressed as mean S.E.M. For the drug-induced rotational behavior, non-parametric Kruskall-Wallis test was used. Inter-group differences for values of Nissl-stained neurons for the injected side and biochemical assays were found out using one-way ANOVA followed by Tukey’s post-hoc test. In all analyses, the null hypothesis was rejected at a level of 0.05.