Bacterial strain and plasmid
E. coli BL21 F-
ompT hsdSB (rB-mB-)
gal dcm (DE3) (Invitrogen, CA, USA) was used as the host for synthesis of rhINF-β. An oligonucleotide was designed regarding
E. coli codon usage; using vectorNTI version 11.0 (Invitrogen, CA, USA) (
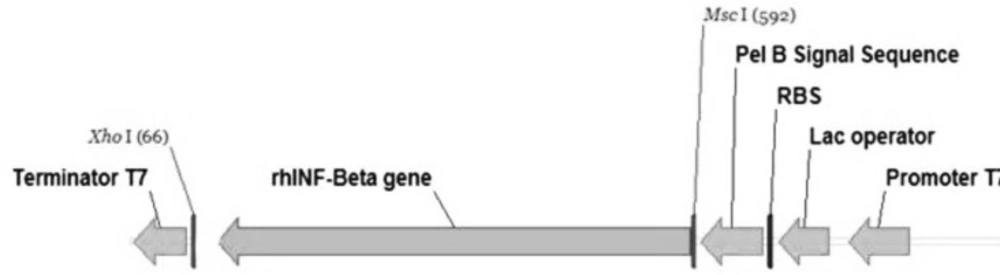
26) and synthesized. The in-house software optimizes a variety of parameters that are critical to the efficiency of gene expression, including codon usage bias, GC content, CpG dinucleotides content, mRNA secondary structure, cryptic splicing sites, premature polyA sites, internal Chi (crossover hot-spot instigator) sites and ribosomal binding sites, negative CpG islands, RNA instability motifs, AU-rich elements, repeat sequences (direct repeats, reverse repeats, and dyad repeats or palindromes), and restriction sites that may interfere with cloning procedure. It was inserted into the
XhoI and
MscI cloning sites. The pET-25b(+) vector (Novagen, CA, USA) containing the requested insert including the optimized synthetic human INF-β-1b was purchased from Biomatik Corporation, ON, Canada.
Briefly, the designed plasmid consists of the strong inducible T7 promoter under the control of the lac-operator sequence, the ORF for encoding gene of the requested protein, followed by the T7 transcription terminator transcription, a ColE1 origin of replication, the β-lactamase gene under its own promoter to confer resistance to ampicillin with the strong transcription terminator of phage λ.
The prokaryotic host cells were transformed using CaCl2 method with the constructed plasmid and transforming clones were selected on LB (Luria-Bertani) medium with 100 μg mL-1 ampicillin. Plasmids were prepared by the alkaline lysis method (
27) and purified using plasmid kits from Vivantis (Selangor, Malaysia).
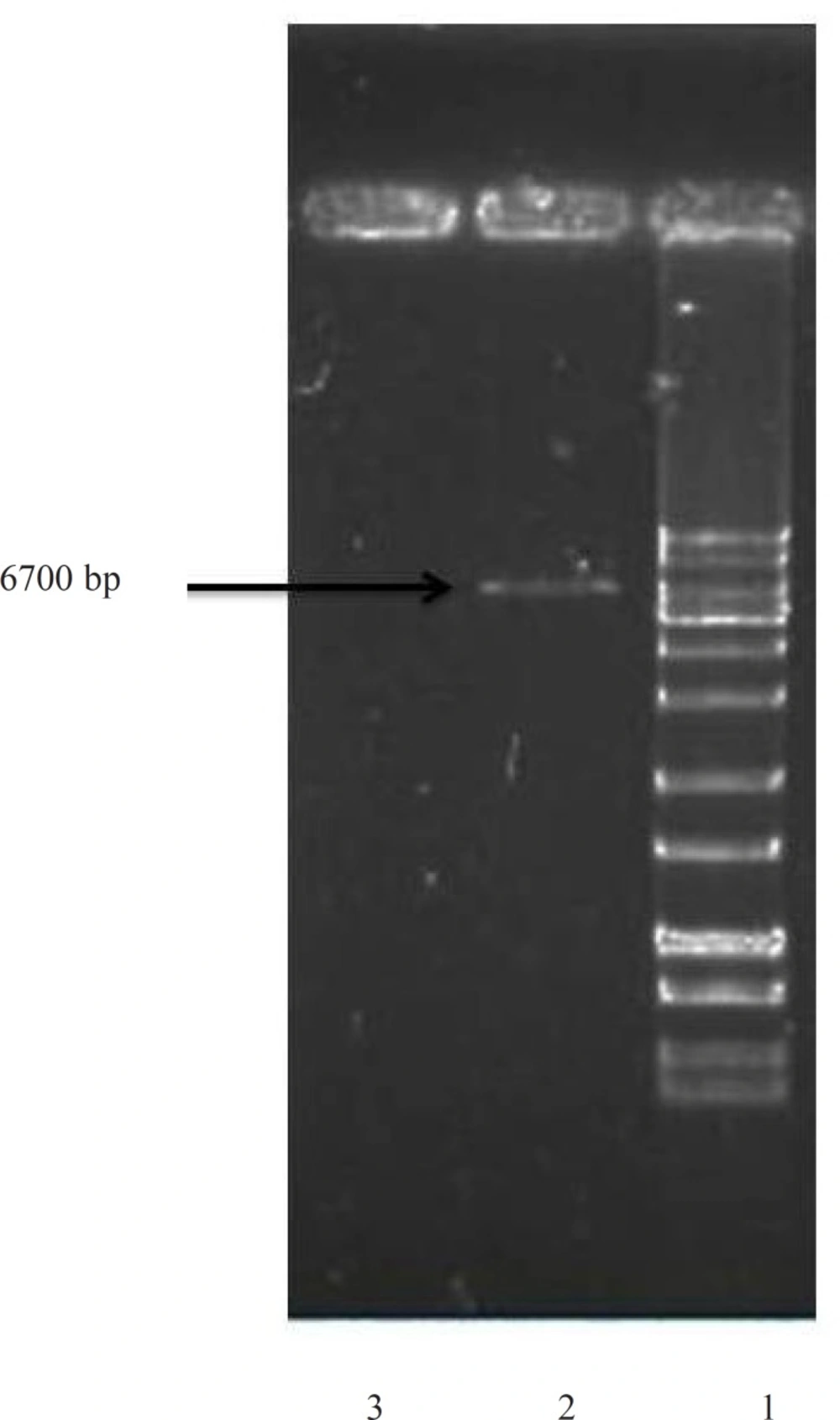
Cloning into plasmid vectors was performed by standard methods (
28). The insertion was sequenced to confirm its integrity. The bacterial stocks were kept at −70 °C in 20 % (v/v) glycerol for long-term usage.
Shake flask culture
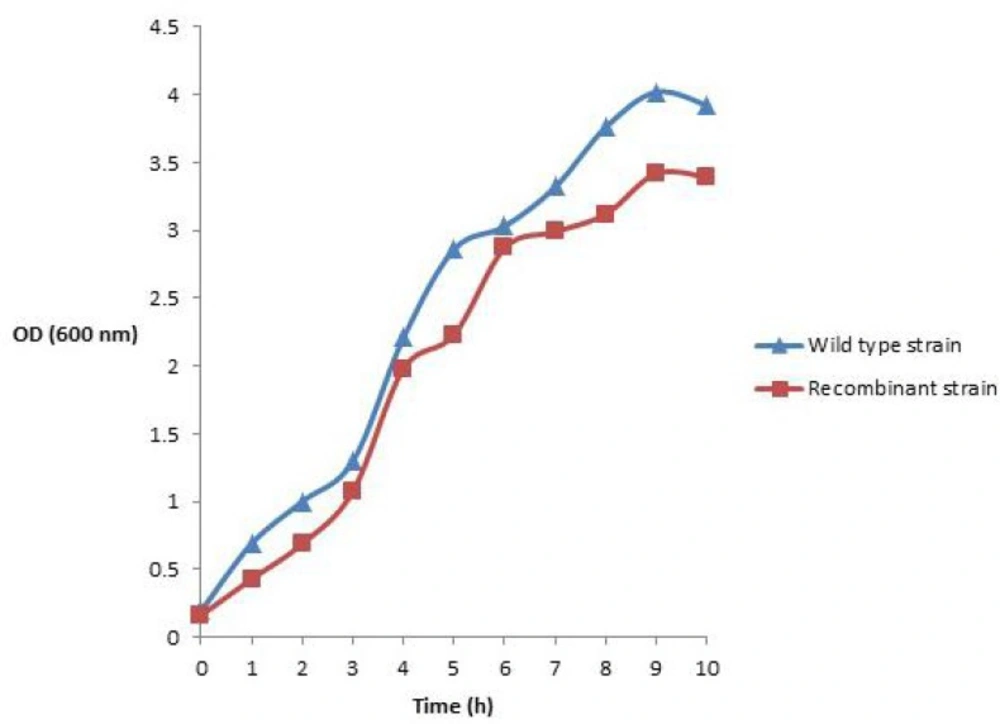
A frozen culture (1.5 mL) of recombinant transformed cells was inoculated into 50 mL of primary culture medium (sterile TB medium) containing 100 μg mL-1 of ampicillin in a 250 mL Erlenmeyer shake flask and incubated overnight in an NB-205V shaking incubator (N-BIOTEK, Bucheon-si, South Korea) at 160 rpm, 37 °C. Primary seed culture (1 mL) was used to inoculate 100 mL of secondary culture medium (sterile corresponding medium) in a 500 mL Erlenmeyer shake flask and grown for 8 h at 37 °C at 160 rpm. The cell growth was monitored during cultivation by optical density measurements at 600 nm using photometry method and the sampling was carried out every one hours.
Inoculum preparation for fermentation
The recombinant plasmid was transformed into E. coli strain BL21 (DE3). An individual colony was selected from LB agar plate and inoculated into 5 mL of terrific broth (TB) medium containing 100 μg mL-1 ampicillin. The 5 mL E. coli culture was incubated in shaking incubator overnight at 37 °C. One hundred μL of the overnight culture was inoculated into 200 mL TB medium containing 100 μg mL-1 of ampicillin. It was grown in shaking incubator at 37 °C and after reaching to OD600nm=0.7-1.0, then it was transferred to the bench-top bioreactor as seed culture.
Batch-culture experiment
The batch fermentation was carried out in a bench-top 5.0 L Minifors fermenter system (Infors HT, Basel, Switzerland) with a working volume of 2 L. The system was equipped with pH, temperature and pO2 probes and four peristaltic pumps for the addition of alkali, acid, nutrients, salts or antifoam solutions. All the fermentation parameters were registered using a computer connection to the fermenter, equipped with IRIS software. It could remotely control all the fermentation parameters, included pH, pO2, agitation speed (rpm) and aeration (L·min-1) and also was used for data storage.
The inlet airflow used was 1.0 vvm. The dissolved oxygen was monitored using a polarographic oxygen electrode (Mettler Toledo, Switzerland) and maintained above 30% saturation throughout the experiment. The pH was measured using a glass electrode and controlled at pH 7.0 using 1.0 M NaOH and 1.0 M HCl. The temperature and agitation were maintained at 37 °C and 500 rpm, respectively. The foam was controlled by the addition of a silicone-based anti-foaming reagent. Samples were taken at different times and analyzed for OD600nm, and rhINF-β production. TB medium was used for the fermentation study and the cell growth was monitored during cultivation by optical density measurements at 600 nm using photometry method.
Induction and expression of rhINF-β
The cells were induced at OD600nm of 1.0 with 0.2 mM IPTG (Fermentas, Vilnius, Lithuania). A part of the culture was used as negative control without adding IPTG (pre-induction). After 1-4 hours of additional growth at 37 °C, cells were harvested by centrifugation at 5000 g for 10 min at 4 °C.
Preparation of periplasmic protein
The pellet was thawed to room temperature and resuspended to 1 mL in a freshly prepared hypertonic solution of cell lysis buffer containing 20 % (w/v) sucrose, 30 mM Tris-Cl (pH 8.0) and 1 mM EDTA (pH 8.0). The mixture was stored on ice for 30 minutes at 4 °C. Cells were centrifuged at 5000 g for 10 min at 4 °C and the supernatant was collected. Cells were resuspended to 1 mL in a hypotonic solution of ice cold 5 mM MgSO4 and incubated for 30 min at 4 °C. The mixture was then centrifuged and the supernatant of hypotonic solution was collected and combined with the supernatant from the hypertonic solution, followed by an additional centrifugation to remove debris. The supernatant was collected immediately for the periplasmic proteins. The total soluble protein was analyzed by the Bradford method with bovine serum albumin (BSA) as a standard (
29).
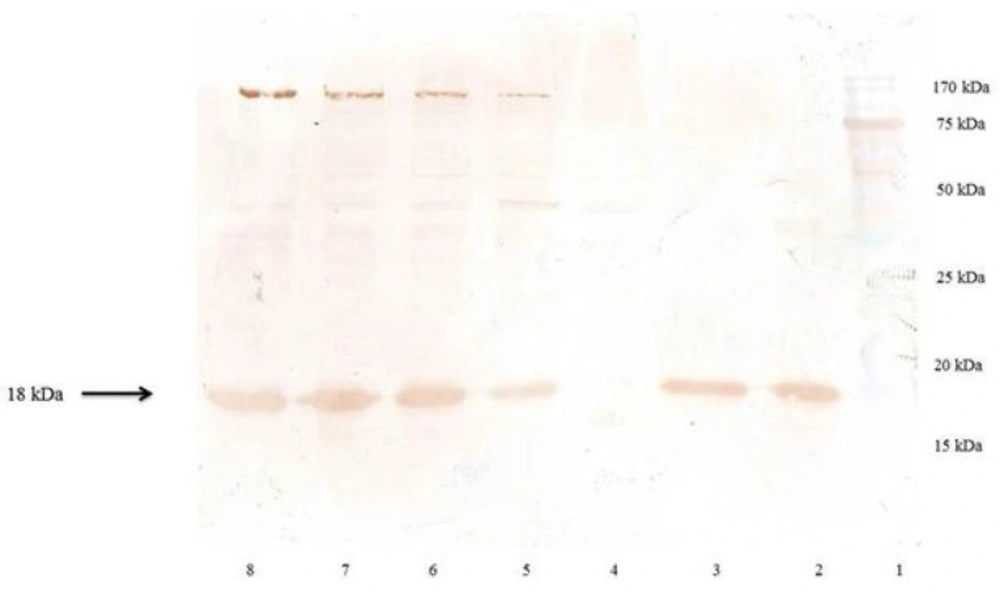
Immunoblot analysis of the Recombinant hINF-β
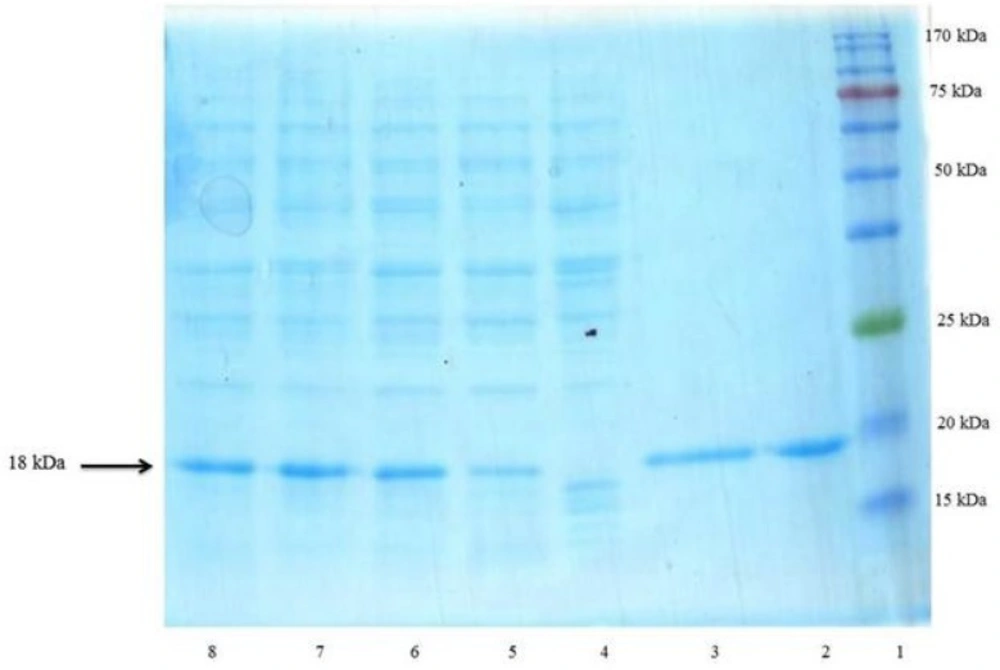
The resulting bacterial pellet or purified protein were homogenized in SDS sample loading buffer containing 0.150 M Tris–Cl, pH 6.8; 10 % glycerol; 2 % SDS; 0.01 % bromophenol blue; and 0.5 M 2-ME (2-Mercaptoethanol). It was mixed properly and boiled for 10 min and centrifuged at 11,000 g for 2 min. Electrophoresis was performed in the presence of SDS according to the method of Laemmli (
30). The discontinuous gel consisted of a 4 % stacking gel and a 17.5 % separating gel was run on a vertical electrophoresis unit (omniPAGE mini, Cleaver Scientific, UK). Two commercial non-glycosylated rhINF-βs (Betaseron®, Bayer HealthCare, Germany and ZiferonTM, Zist Daru Danesh, Tehran, Iran) were used as the standard. Gels were stained with Coomassie Brilliant Blue R250. The total protein pattern of the recombinant bacteria, visualized on Coomassie brilliant blue stained gel, were scanned by a Bio-Rad Gel Doc 2000 densitometric gel scanner with an accuracy of greater than 95 %.
For western blotting experiment, electrophoresed proteins were transferred to a nitrocellulose membrane using a semi-dry method in a transfer buffer (25 mM Tris, 192 mM glycine, 20 % methanol) at 20 mA for 7 minutes using iBlot® western detection kit (Invitrogen, CA, USA). The transferred membrane was incubated firstly with 2 % bovine serum albumin (BSA) in phosphate buffered saline/Tween 20 (PBST) for one hour, followed by primary antibody (Anti-β-IFN Mouse mAb, Calbiochem, Merck Millipore, Darmstadt, Germany) in 1 % BSA/PBST for one hour and then with secondary antibody (Goat Anti-Mouse Total Ig Peroxidase conjugate, Calbiochem, Merck Millipore, Darmstadt, Germany) in 1 % BSA/PBST for one hour. The membrane was washed with PBST for three times after each incubation period. The blot was then developed using diaminobenzidine (DAB) tetrahydrochloride and hydrogen peroxide.
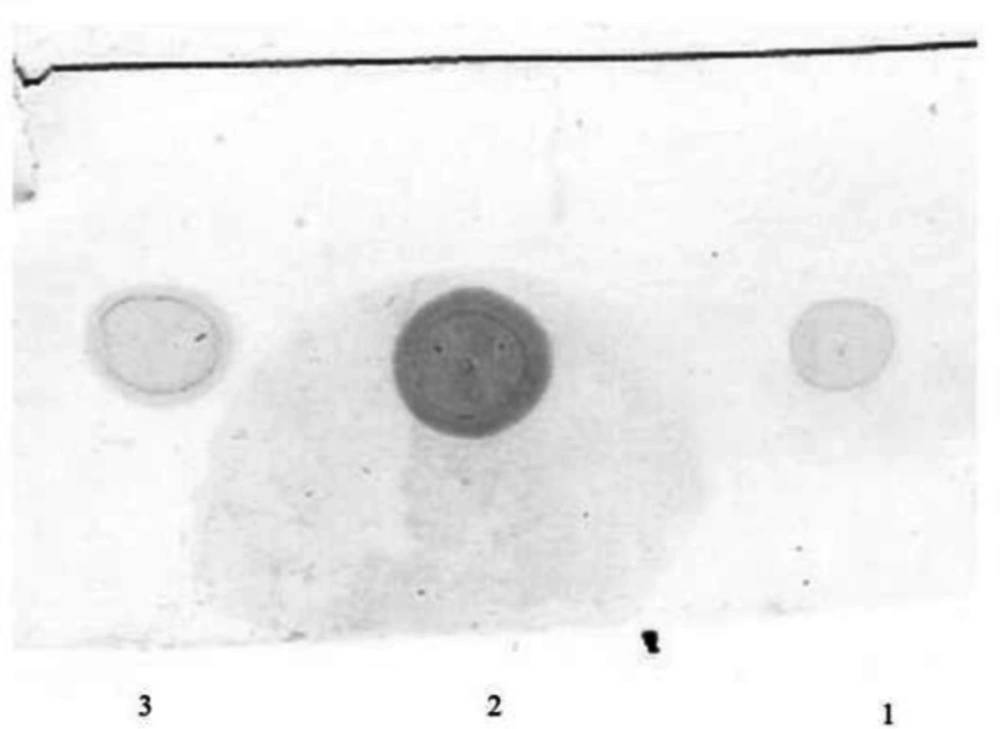
Dot blot
1.5 μL of the extracted recombinant hINF-β was applied to a nitrocellulose membrane and allowed to dry. The membrane was incubated for one hour at 4 °C in a blocking solution consisting of 2 % BSA in TBS-T pH 7.5. The membranes were washed three times for 10 min with TBS-T and incubated for one hour with Anti-β-IFN Mouse mAb (Calbiochem, Merck Millipore, Darmstadt, Germany) diluted 1:500 in TBS-T. The membranes were washed three times for 10 min with TBS-T and then, after addition of the secondary antibody, the rhINF-β spots were revealed. After color development, the membranes were washed out with water and the color changing was evaluated in both transformed and wild strains.