Materials
rhIFNα2b was kindly gifted by Pasteur Institute of Iran (Tehran, Iran). Acrylamide/bis acrylamide solution, ammonium peroxodisulfate, bromophenol blue sodium salt, Coomassie brilliant blue, 1,4-dithiothreitol (DTT), glycerol, glycine, iso-butanol, 2-mercaptoethanol, sodium chloride, sodium dodecyl sulfate (SDS), di-sodium hydrogen phosphate anhydrous, sodium dihydrogen phosphate 1-hydrate, tetramethylethylenediamine (TEMED), and tris (hydroxyl methyl) amino methane (Tris) were purchased from Merck (Darmstadt, Germany). All solutions were prepared with sterile deionized water (Millipore Company, USA). Buffers were filtered through 0.2 μm membranes prior to use.
Stress studies
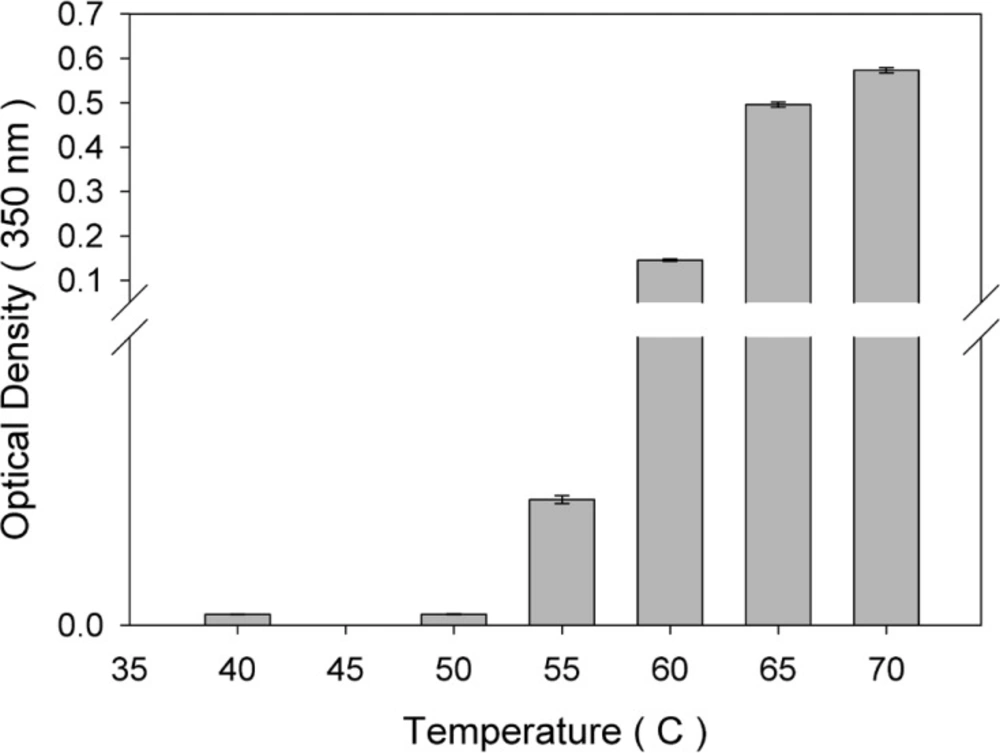
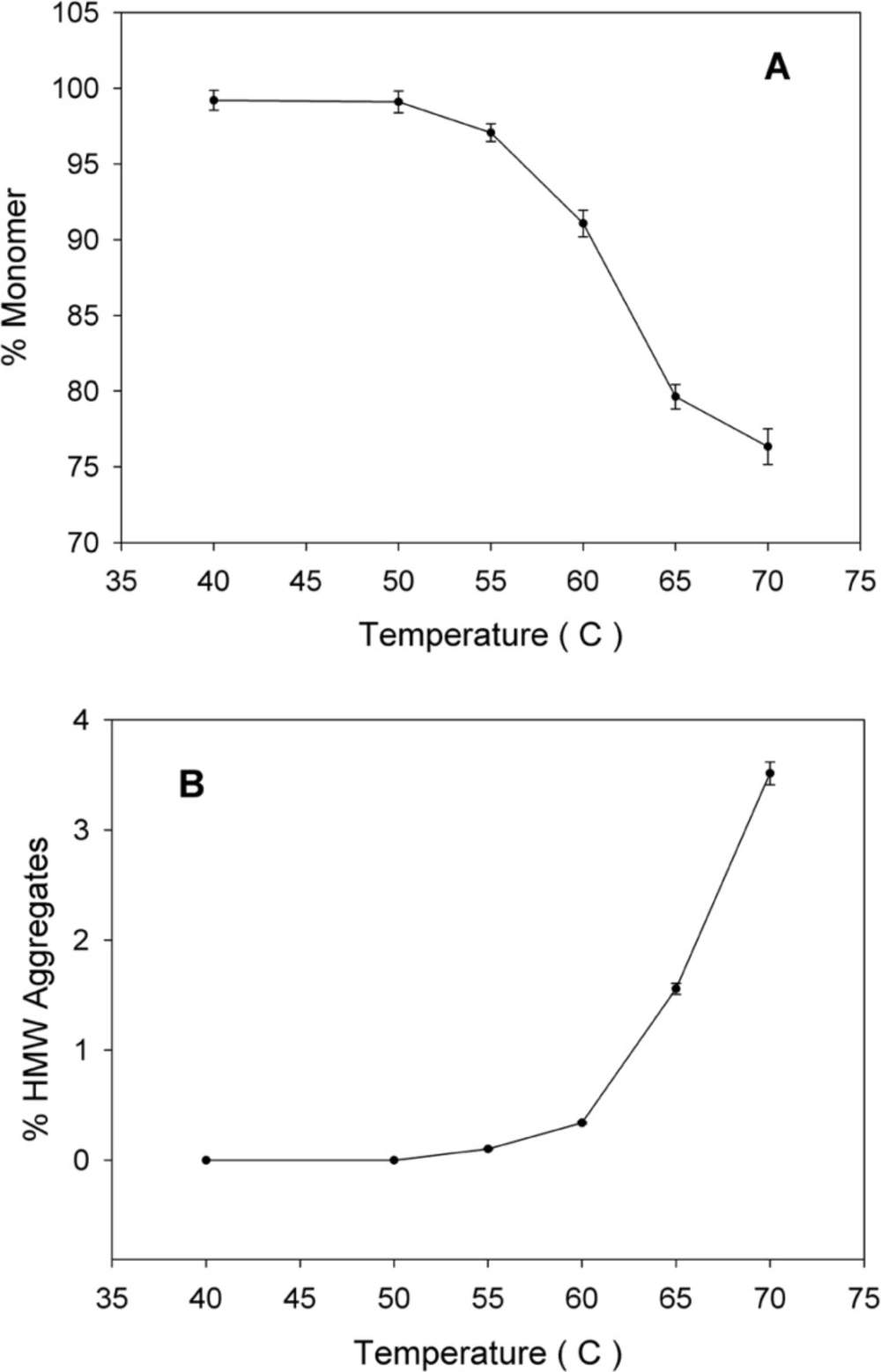
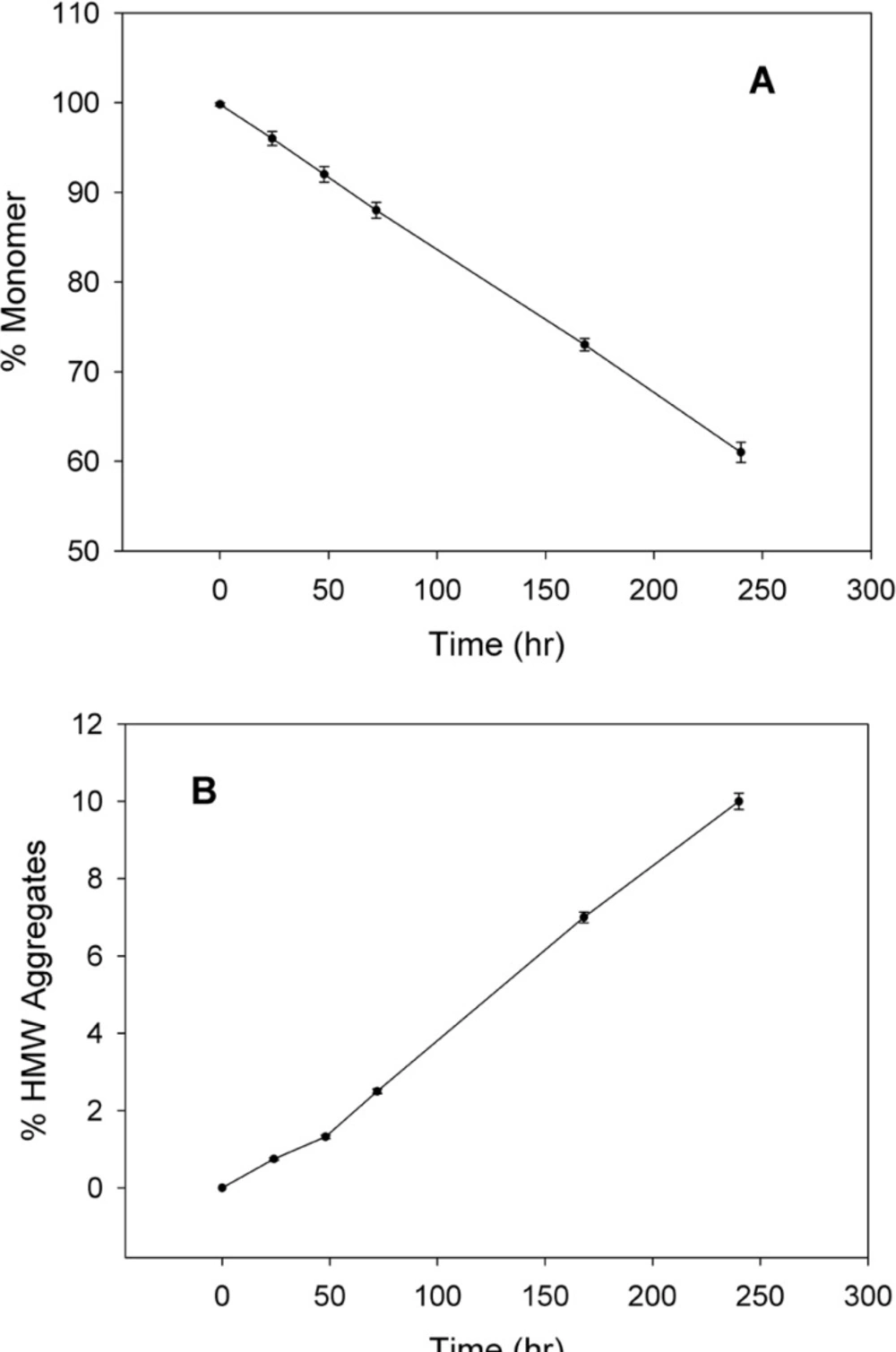
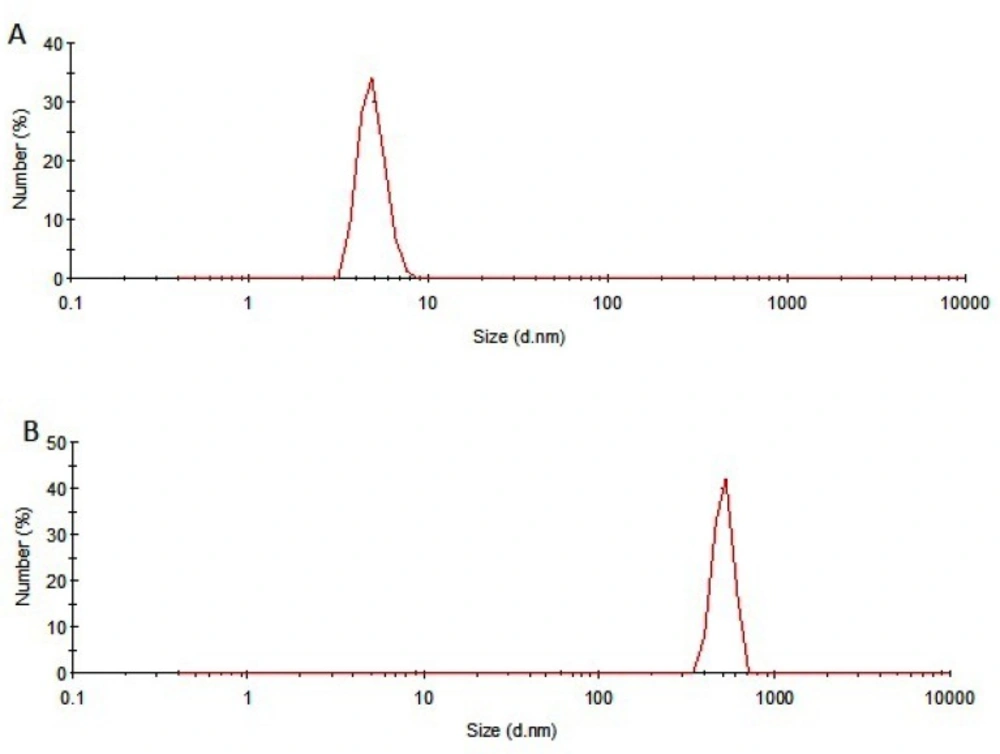
The thermal stability of rhIFNα2b was investigated following heating the protein samples. Protein samples composed of 100 μg/mL in 100 mM sodium phosphate buffer solution (pH 7.0) were incubated (IFE 500, Memmert, Germany) for 1 h at various temperatures (40, 50, 55, 60, 65 and 70 °C) and then cooled to room temperature. The level of heat-induced aggregation and monomer content were monitored by both turbidity measurement (optical density determination) at 350 nm and size exclusion high performance liquid chromatography (SE-HPLC). In another experiment, the samples were incubated at 50 °C (up to 240 h) and analyzed by SE-HPLC, optical density determination, dynamic light scattering and electrophoresis techniques. The relationship between thermal stability of protein with pH of sodium phosphate buffer solution (
5,
6, and
7) and buffer concentrations (10, 55, and 100 mM) were also studied through heating the protein samples (100 μg/mL) at 50 °C for 72 h by SE-HPLC.
Turbidity measurements
Optical density of the protein sample was determined by ScanDrop® 250 (Analytik Jena AG, Jena, Germany) at 350 nm against sodium phosphate buffer as the blank solution to evaluate the aggregate production.
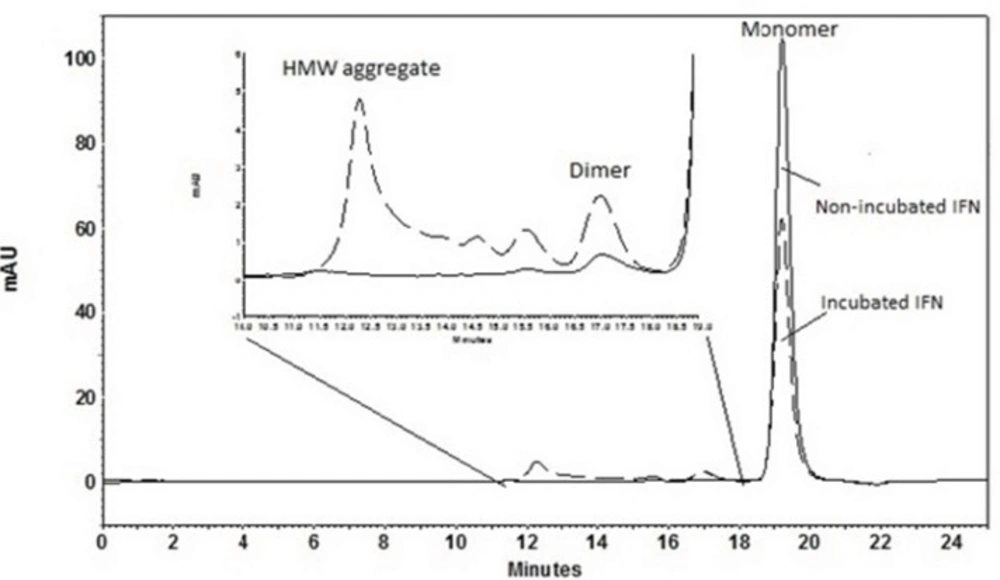
Size exclusion high performance liquid chromatography (SE-HPLC)
Size exclusion separation was carried out on a TSKgel G3000 SWXL column (7.8 mm ID×300 mm, 5-μm particles, 250Å pore size, Tosoh Bioscience, Japan). Analysis was performed at 25 °C under isocratic elution at a rate of 0.5 mL/min with a mobile phase consisting of 100 mM sodium phosphate (pH 7.0), 150 mM NaCl and 0.05% (w/v) sodium dodecyl sulfate (SDS).The column was calibrated by protein standard markers.
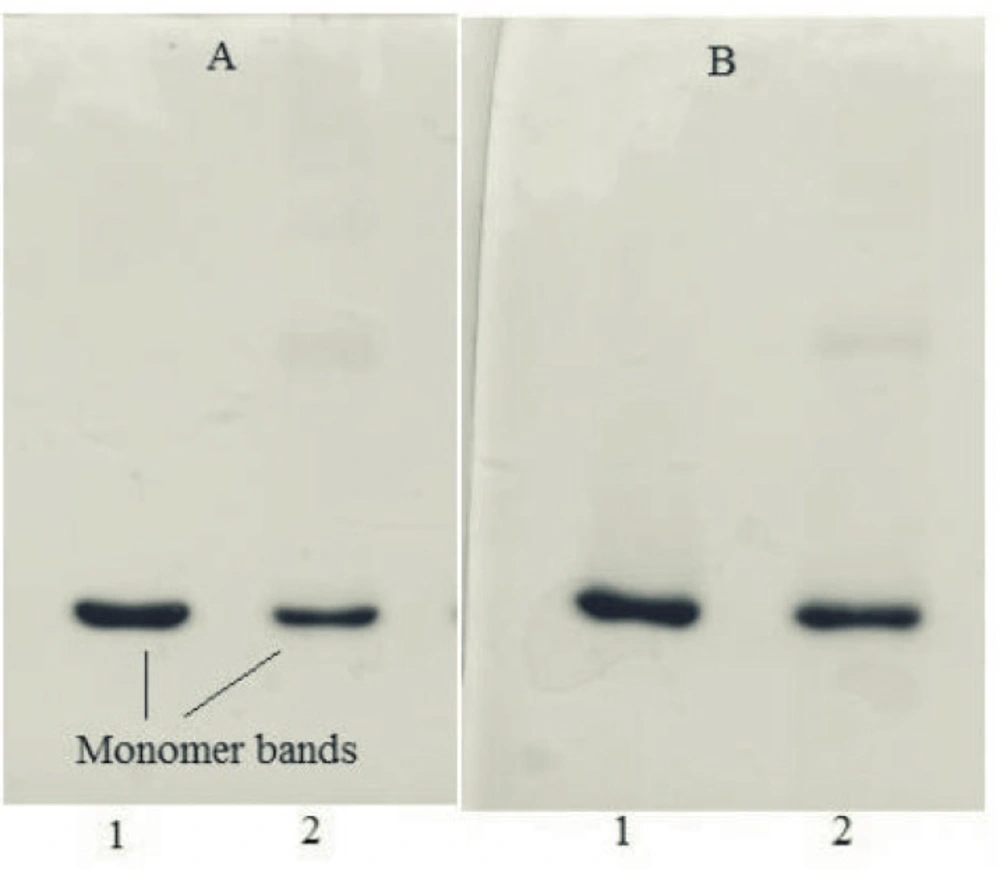
Sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE)
Gels were composed of a separating gel containing 12.5% (w/v) acrylamide and 0.1% (w/v) SDS, and a stacking gel containing 3% (w/v) acrylamide and 0.1% (w/v) SDS. Gels were run under reducing (sample buffer containing 5% v/v β-mercaptoethanol) and non-reducing (sample buffer without β-mercaptoethanol) conditions at 200 V at room temperature, using a MP300 electrophoresis instrument (Cleaver Scientific, UK).
Dynamic light scattering (DLS)
A Malvern Zetasizer Nano ZS (Worcestershire, UK) laser light scattering system, equipped with a Nano ZS® software for data acquisition and analysis, was used for particle size measurements.
Statistical analysis
Results are reported as mean ± SD. Data obtained was compared using one-way ANOVA. Differences between the treatments were assumed to be significant at p < 0.05. Statistical analysis of all data was performed using IBM® SPSS® Statistics, v19, 2010.