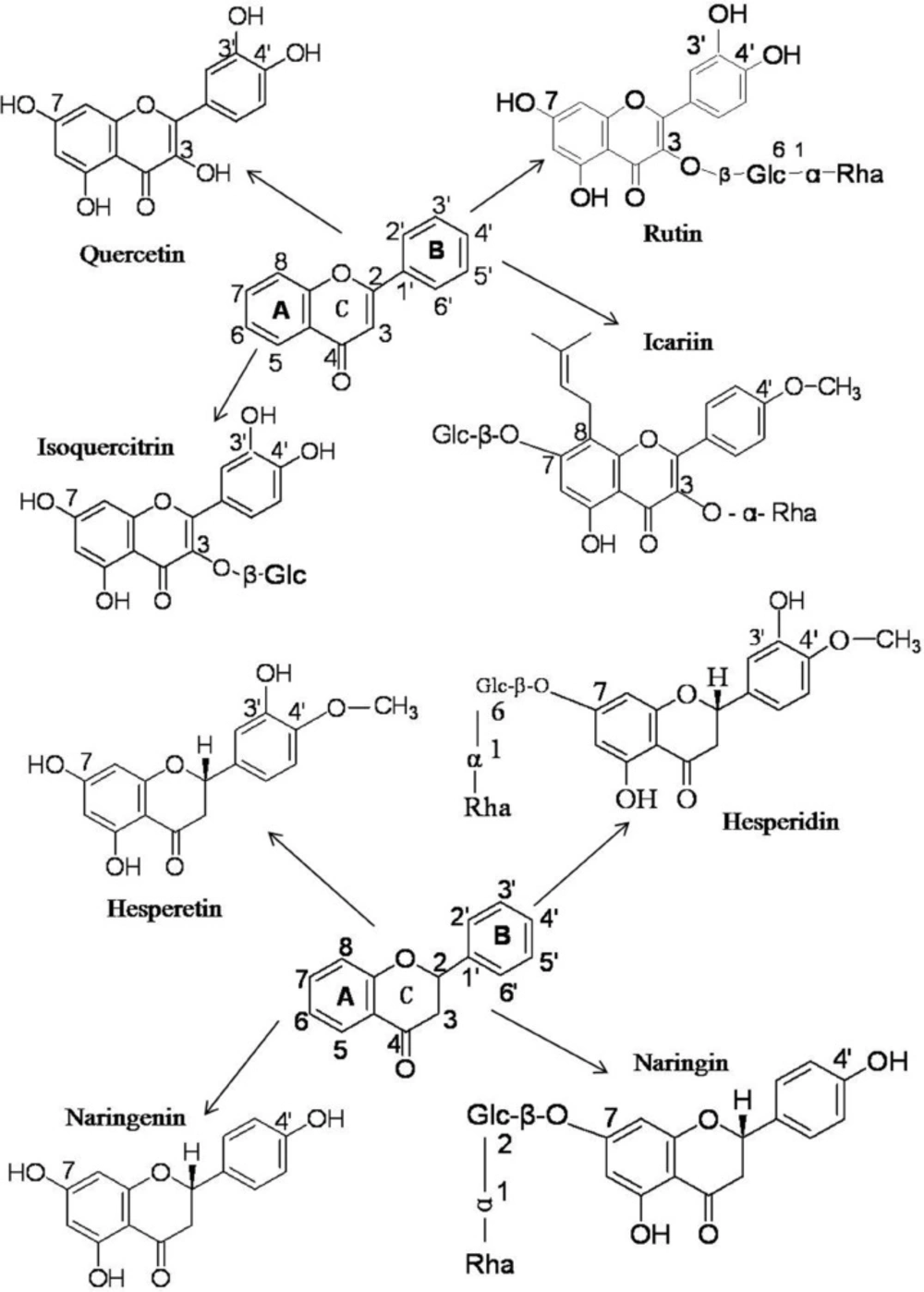
Structures of flavonoid phytochemicals
The flavonoids utilized the current work can be cataloged as flavonols (quercetin), and flavones (rutin, isoquercitrin and icariin), which all share the core structure of flavone and flavanones (hesperetin, hesperidin, naringenin and naringin) (
Figure 1). On one hand, these flavonoids share a similar structure, but on the other hand they also have their individual intrinsic structures (
Figure 1). The structural differences among these flavonoids might result in and explain their functional differences.
The chemical structures of quercetin, rutin, isoquercitrin, icariin, hesperetin, hesperidin, naringenin and naringin
Purity analyses of flavonoid phytochemicals
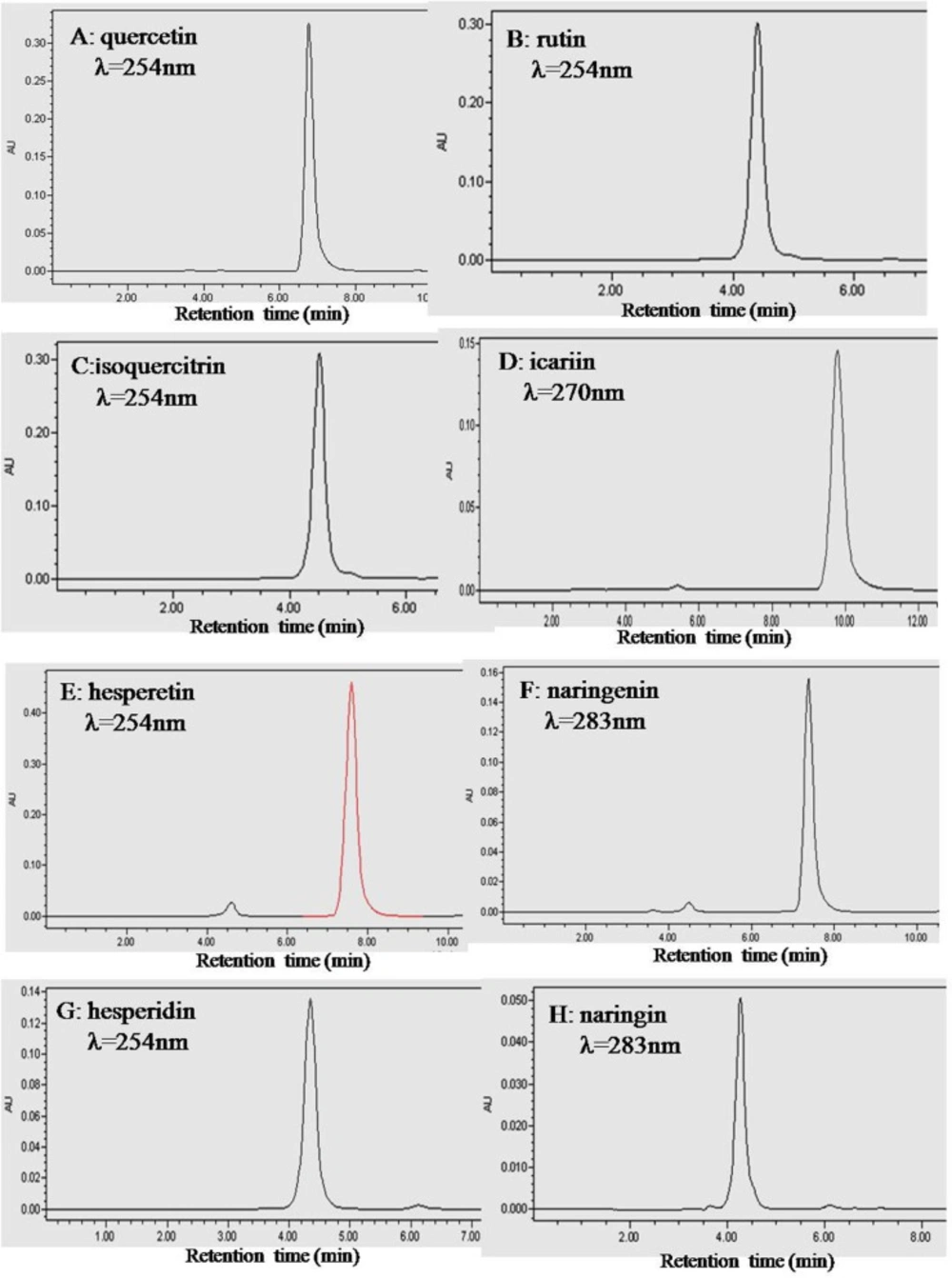
To ensure that the fluorescence results we obtained were indeed due to the flavonoid, a purity analysis was carried out. The HPLC chromatograms of quercetin, rutin, isoquercitrin, icariin, hesperetin, naringenin, hesperidin and naringin are shown in
Figure 2. The peak of each individual flavonoid dominated its chromatogram and the calculated purities of the eight flavonoids were over 95% based on both the peak area and intensity height (
Table 1).
HPLC chromatograms of the eight flavonoids. Plots A, B, C, D, E, F, G and H are for quercetin, rutin, isoquercitrin, icariin, hesperetin, naringenin, hesperidin and naringin, respectively. HPLC was performed with an Intersil ODS-3 C18 column (4.6 × 250 mm) at 25 oC. The mobile phase was 60% (v/v) methanol containing 0.2% phosphoric acid. The column was eluted with a flow rate of 1.0 mL/ min. The absorbance wavelength was set at 254 nm for quercetin, rutin, isoquercitrin, hesperetin and hesperidin, 283 nm for naringenin and naringin, and 270 nm for icariin.
| Flavonoids | Na | Wavelength (nm)b | Retention time (min) | Purity (area) (%)c | Purity (height ) (%)d |
|---|
| Quercetin | 3 | 254 | 6.75 ± 0.01 | 98.22 ± 0.02 | 98.49 ± 0.02 |
| Isoquercitrin | 3 | 254 | 4.51 ± 0.01 | 97.12 ± 0.02 | 97.20 ± 0.02 |
| Rutin | 3 | 254 | 4.40 ± 0.01 | 99.33 ± 0.03 | 99.33 ± 0.02 |
| Naringenin | 3 | 254 | 7.38 ± 0.02 | 95.64 ± 0.02 | 95.10 ± 0.02 |
| Naringin | 3 | 254 | 4.26 ± 0.01 | 95.06 ± 0.04 | 95.31 ± 0.03 |
| Hesperetin | 3 | 283 | 7.83 ± 0.02 | 95.85 ± 0.03 | 95.02 ± 0.03 |
| Hesperidin | 3 | 283 | 4.35 ± 0.02 | 96.53 ± 0.02 | 96.78 ± 0.03 |
| Icariin | 3 | 270 | 9.78 ± 0.02 | 96.85 ± 0.03 | 96.29 ± 0.03 |
N = the number of measurements per experiment;
The wavelength used for monitoring the elution of flavonoid phytochemicals by HPLC;
The purities of eight flavonoid phytochemicals were calculated based on their peak areas;
The purities of eight flavonoid phytochemicals were calculated based on their peak heights.
Binding of flavonoid phytochemicals to BSA
Epidemiological evidence indicates that a diet rich in flavonoids reduces the incidence of chronic diseases such as cardiovascular diseases, diabetes, cancers, neuronal diseases and stroke (
29,
30). However, whether flavonoids play a significant role in preventing such chronic diseases has been questioned because of their low bioavailability (
31). The interactions of flavonoids with SA determine their bioavailability and toxicology. BSA-flavonoid binding has been investigated by analyzing the perturbation of the intrinsic BSA fluorescence due to its Trp residues, which is observed by excitation at a wavelength of 280 nm (
5,
14,
15). The fluorescence emission intensity of BSA has been observed to decrease as flavonoid concentration increases until at a 1:1 BSA:flavonoid ratio the complex is not fluorescent. Thus flavonoid molecules quench the Trp fluorescence, indicating that they bind near the Trp position (
5,
14,
15). Our results indicate that as the flavonoid concentration increases, the fluorescence intensity of BSA decreases remarkably, implying that flavonoids we have studied also bind with BSA at or near the Trp residues.
Figure 3 shows the plots of log ΔI/I
versus log [D] for quercetin, rutin, isoquercitrin, icariin, hesperetin, naringenin, hesperidin and naringin for concentrations, [D], between 0.5 μM and 5 μM.
Double-logarithm curves of BSA fluorescence for flavonoids. A, B, C, D, E, F, G and H are the plots of log (ΔI/I) versus log [D] of quercetin, rutin, isoquercitrin, icariin, hesperetin, naringenin, hesperidin and naringin with the concentration ranges of 5.30×10- 7-5.30×10-6 M, 8.30×10-7-3.73×10-6 M, 1.55×10-6-5.18×10-6 M, 1.04×10-6-5.18×10-6 M, 1.04×10-6-5.18×10-6 M, 5.18×10-7-5.18×10-6 M, 4.15×10-7-3.73×10-6 M and 8.30×10-7-4.15×10-6 M, respectively. Significant differences exist for all comparisons between any two individual flavonoids (P<0.05) by SPSS 13.0 statistical analysis.
The K
A values and the number (n) of binding sites for all eight flavonoids were calculated and are summarized in
Table 2. Hesperetin had the greatest K
A for binding with BSA (5.59×10
5), followed by quercetin (4.94×10
5), naringenin (3.04×10
5), isoquercitrin (4.66×10
4), icariin (3.60×10
4), rutin (1.65×10
4), hesperidin (2.50×10
3) and naringin (8.70×10
2). SPPS 13.0 analysis indicated that the differences of binding activities of all eight flavonoid phytochemicals used in the current work are statistically significant (
P<0.05). For all eight flavonoids the number of BSA binding sites was close to 1 (
Table 2).
The binding capacities of flavonoids with BSA are determined by the specifics of their structures. Rutin and isoquercitrin share the same core flavone structure with quercetin but have no free 3-OH group on the C-ring due to their O-Glc-α-Rha and O-β-Glc substituents (
Figure 1), which leads to a decrease in the BSA binding association constant, K
A, from 4.94 × 10
5 to 4.66 × 10
4 and 1.65 × 10
4, respectively. A similar phenomenon was also observed for hesperetin and hesperidin, and for naringenin and naringin. When the free 7-OH groups in the A-ring of hesperetin and naringenin were replaced by 7-O-β-Glc(6→1)-α-Rha and 7-O-β-Glc(2→1)-α-Rha to become hesperidin and naringin, respectively, their BSA binding association constants decreased from 5.59×10
5 to 2.50×10
3 and from 3.04×10
5 to 8.70×10
2. Considering that the only structural differences between hesperidin and hesperetin, and naringin and naringenin are the replacement of the 7-OH group by 7-O-β-Glc (6→1)-α-Rha and 7-O-β-Glc(2→1)-α-Rha, it is apparent that the presence of the free 7-OH enhances the binding capacities of flavanones with BSA. The hydroxyl group can interact with amino acid residues of BSA to form hydrogen bonds to improve the binding affinities. When the free -OH is replaced by saccharides, the molecular size increases and the resultant steric hindrances in the binding pocket result in a lower binding affinity to BSA. Furthermore, the additional polar saccharide groups render these molecules less hydrophobic, and could affect their orientation in relation to the hydrophobic environment of BSA (
32-
38). Thus substitution of the 3-OH and 7-OH groups has marked effects on the binding affinities of flavonoids with BSA.
When the 3-OH in the C-ring of quercetin was replaced by 3-O-Glc-α-Rha in rutin and 3-O-β-Glc in isoquercitrin, the binding affinities to BSA decreased by one order of magnitude, but when the 7-OH groups in the A-rings of hesperetin and naringenin were replaced by 7-O-β-Glc(6→1)-α-Rha and 7-O-β-Glc(2→1)-α-Rha in hesperidin and naringin, the binding affinities dropped by two orders of magnitude.
The result indicates that substituents on the hydroxyl group of C7 in the A ring are more important for flavonoid interaction with BSA than are substituents on the hydroxyl group of C3 in the C-ring and thus that the A ring might be the more critical structure for binding to the hydrophobic region of BSA.
In comparison, the binding affinity of icariin with BSA was much higher than of hesperidin and naringin. The difference in affinity can be ascribed to the 7-O-β-Glc in the A ring of icariin being smaller than the 7-O-Glc-α-Rha of hesperidin and naringin. In addition, the 8-alkyne of icariin enhances its lipid solubility and lipid-soluble drugs are more likely to enter the hydrophobic cavity of BSA and form a drug- BSA complex (
33,
34,
36,
37). The two effects combine to contribute to the higher binding affinity of icariin with BSA.
Taken together, the current work indicates that the binding capacities of flavonoid phytochemicals with BSA are related to their structures. The above results agree with published data for the binding affinities of other kinds of flavonoids with BSA, for which hydrogen bonding, hydrophobic forces and steric hindrance effects have been considered to be the driving forces for BSA-flavonoid association (
3,
9,
14,
27,
28,
32-
38). The study of the interactions of quercetin, rutin, hyperin and baicalin with HSA, indicated that flavonoids containing the groups C(3)–OH and C(4)=O, or C(5)–OH and C(4)=O, or C(3′)–OH and C(4′)– OH could form flavonoid-HSA complexes (
9,
14,
28,
32). The numbers of hydroxyl groups on the A and B-rings of flavonoids affected the binding affinities and the number of binding sites of flavonoids with BSA significantly. Increasing numbers of hydroxyl groups on the A- and B-rings increased binding affinities of flavonols with BSA, presumably by forming hydrogen bonds. BSA has hydrophobic groups in the interior of its tertiary structure and polar groups at its surface. Hydrogen bonds may take place between –OH groups of flavonoids and polar groups at the BSA surface (
35,
36). Glycosylation of their hydroxyl groups decreased the binding affinities of flavonoids with proteins (
37,
38), as confirmed by our results. Glycosylation increases the polarity of the molecule and this lessens the ability of the flavonoid to penetrate into the Trp-rich hydrophobic interior regions of BSA. Glycosylation also increases the size of the molecule and steric hindrance effects in the binding pocket may also weaken the binding affinity of the flavonoid with BSA. Finally, glycosylation decreases the hydrophobicity of flavonoids, reducing their binding affinity with BSA (
37,
38).

![Double-logarithm curves of BSA fluorescence for flavonoids. A, B, C, D, E, F, G and H are the plots of log (ΔI/I) <i>versus </i>log [D] of quercetin, rutin, isoquercitrin, icariin, hesperetin, naringenin, hesperidin and naringin with the concentration ranges of 5.30×10<sup>- 7</sup>-5.30×10<sup>-6</sup> M, 8.30×10<sup>-7</sup>-3.73×10<sup>-6</sup> M, 1.55×10<sup>-6</sup>-5.18×10<sup>-6</sup> M, 1.04×10<sup>-6</sup>-5.18×10<sup>-6</sup> M, 1.04×10<sup>-6</sup>-5.18×10<sup>-6</sup> M, 5.18×10<sup>-7</sup>-5.18×10<sup>-6</sup> M, 4.15×10<sup>-7</sup>-3.73×10<sup>-6</sup> M and 8.30×10<sup>-7</sup>-4.15×10<sup>-6</sup> M, respectively. Significant differences exist for all comparisons between any two individual flavonoids (P<0.05) by SPSS 13.0 statistical analysis.](https://brieflands.com/journals/ijpr/articles/125512/figures/ijpr-13-1019-g003-preview.webp)
