Preparation for surgery and drug treatment
Male Sprague–Dawley rats (200 g to 250 g) were intraperitoneally anesthetized with pentobarbital (70 mg/Kg), intubated, and connected to a respirator. The heart was exposed using an angular incision on the left side of the thoracic cavity. The pericardium was opened to ligate the left anterior descending branch of the coronary artery. Then, the thoracic cavity was closed. After myocardial infarction, the experimental animals were divided into three groups: one group received DOX (25 mg/Kg/day, dissolved in deionized water) twice daily via intraperitoneal injection; one group received MP (50 mg/Kg/day) once a day via intraperitoneal injection and one group were fed a placebo twice daily (the deionized water was the same quantity as that of DOX). The dosages of DOX and of MP are the quantities that affect left ventricular myocardial infarction as reported in the literature (
14). In addition, the sham operation group was subjected to thoracotomy using the same procedures above without ligation of the coronary artery.
Study of cardiac electrophysiology
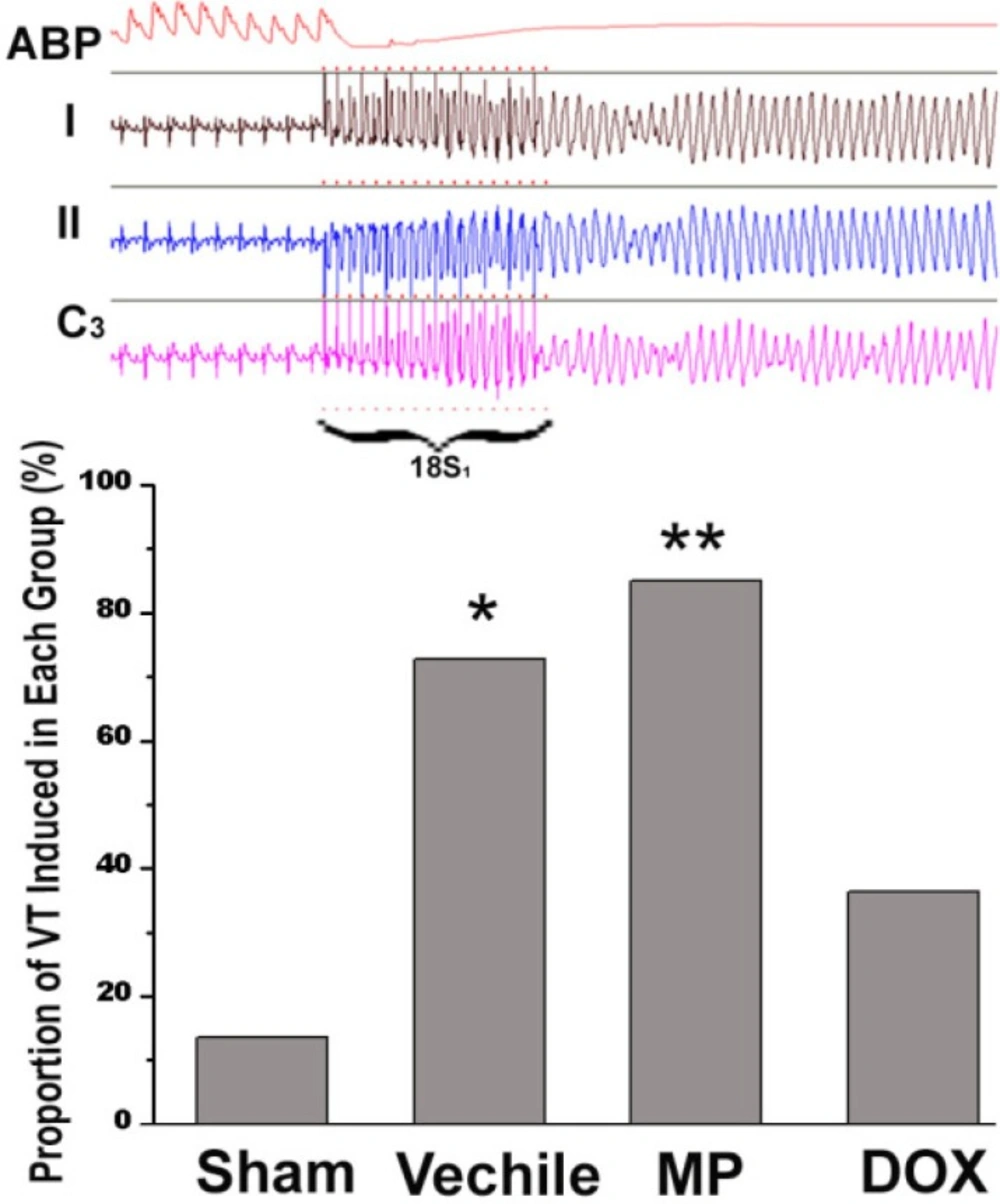
Two weeks after surgery, the rats were anesthetized again, intubated, and connected to a respirator. The chest wall was opened to expose the heart. A customized bipolar electrode was placed on the apex of the right ventricle and attached to the epicardium for pacing. S1S2 stimulation with 130 ms of basic RR interval was used to determine the effective refractory period. Premature stimuli were started from 80 ms and were reduced by 5 ms each time until 20 ms. the burst stimulus (18S1s) was adopted when ventricular tachycardia was not triggered by the S1S2 stimulus. The stimulus interval was steadily increased from 20 ms, ascending by 5 ms each time until a 1:1 capture was formed. Each process was repeated three times. The occurrence of a ventricular tachycardia was monitored continuously during the experiment. A VT duration of >3 s was defined as sustained ventricular tachycardia. Statistical data was collected by replaying video. However, the observers blinded to the grouping of the subjects.
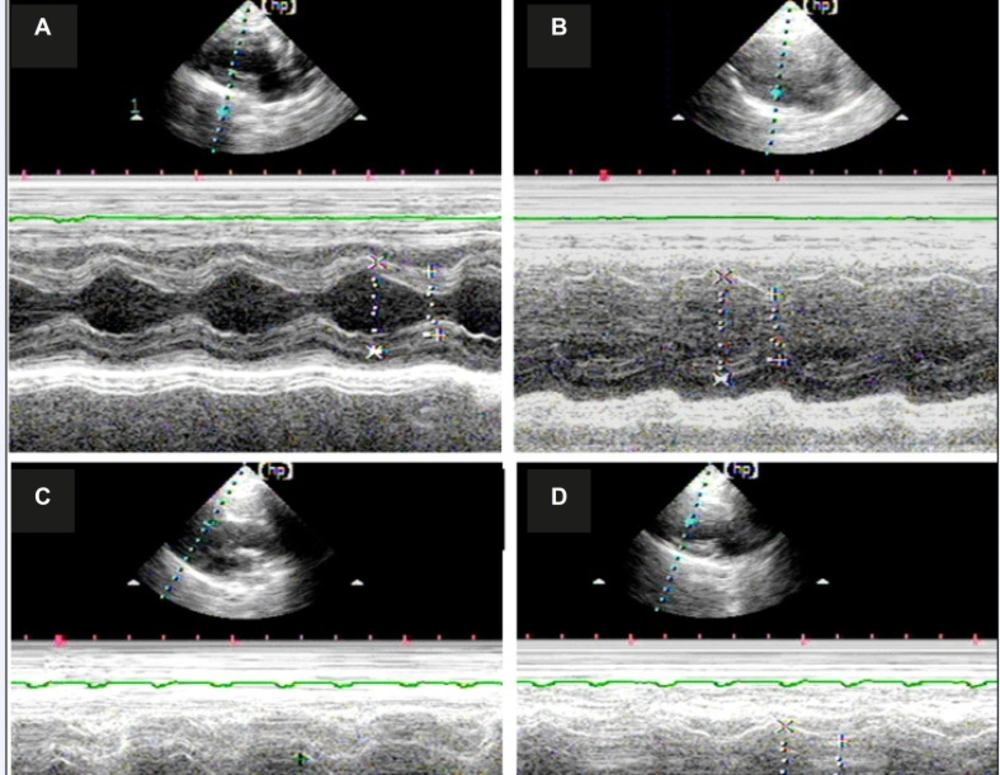
Morphological and functional analysis of the rat heart through echocardiography
Echocardiography study were performed on six animals from each group (n=6) before the electrophysiology study. Two testers blinded to the test grouping conducted the study using an HP5500 cardiac ultrasound machine. The sampling line obtained an M-type ultrasound image passing through the right ventricular anterior wall, the nose of the bicuspid valve, and the posterior left ventricular wall after a 15 MHz probe was used to acquire the typical left ventricular long axis view in two dimensions. The diastolic and systolic left ventricular internal diameters were measured. As a measure of left ventricular (LV) function, the EF (ejection fraction) and fractional shortening (FS) were calculated.
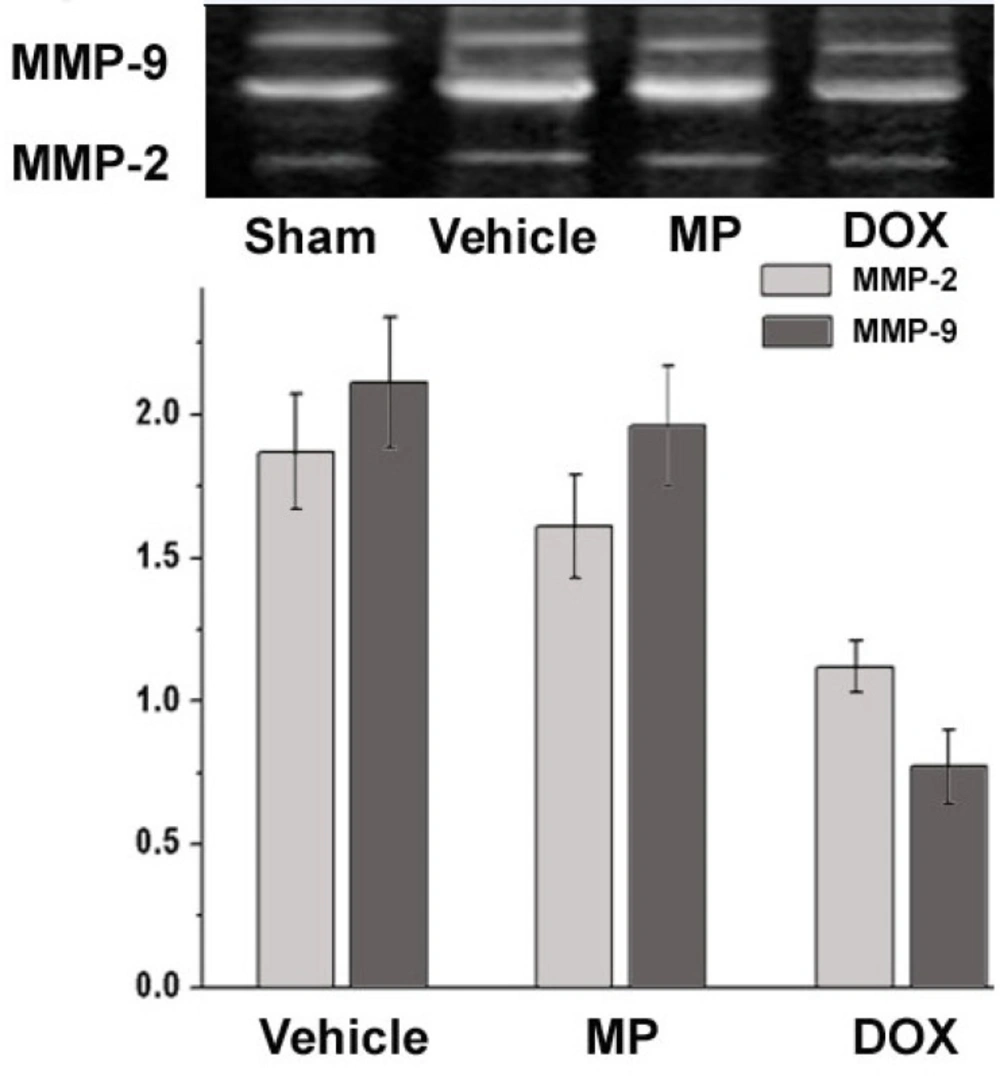
Fluorescence analysis of MMP-2 and MMP-9 activity MMP Zymography
The hearts of the rats were removed after the electrophysiology study. Proteins were extracted from the frozen ventricle (n=6) after homogenization (
16,
17). Briefly, the homogenates were resuspended in lysis buffer, centrifuged at 12000_×
g, and protein concentration was determined using the Bradford technique. The samples were diluted in sample buffer [125 mM Tris–HCl (pH 6.8), 20% glycerol, 4% SDS, and 0.04% bromophenol blue] and aliquots with a final protein content of 5 mg were separated by 10% SDS-polyacrylamide electrophoresis (SDS-PAGE) on gels containing 0.6% gelatin. After SDS-PAGE, the gels were washed twice in extraction buffer [20 mM Tris–HCl (pH 7.5), 2 mM CaCl
2, 1 mM ZnCl
2, 0.02% NaN
3, and 2.5% Triton X-100] for 30 min each time and rinsed in water. The gels were incubated overnight at 37 °C in incubation buffer [20 mM Tris–HCl (pH 7.5), 2 mM CaCl
2, and 1 mM ZnCl
2], stained with Coomassie Blue staining solution (0.5% Coomassie R250, 30% methanol, and10% acetic acid) for 2 h, and then destained in distilled deionized H
2O. Imaging software was used to analyze the grey value after the inverse conversion of the gelatin strip. The result was compared with that of the sham operation group and the relative values were used to represent the relative activities of MMP-2 and MMP-9 for each group.
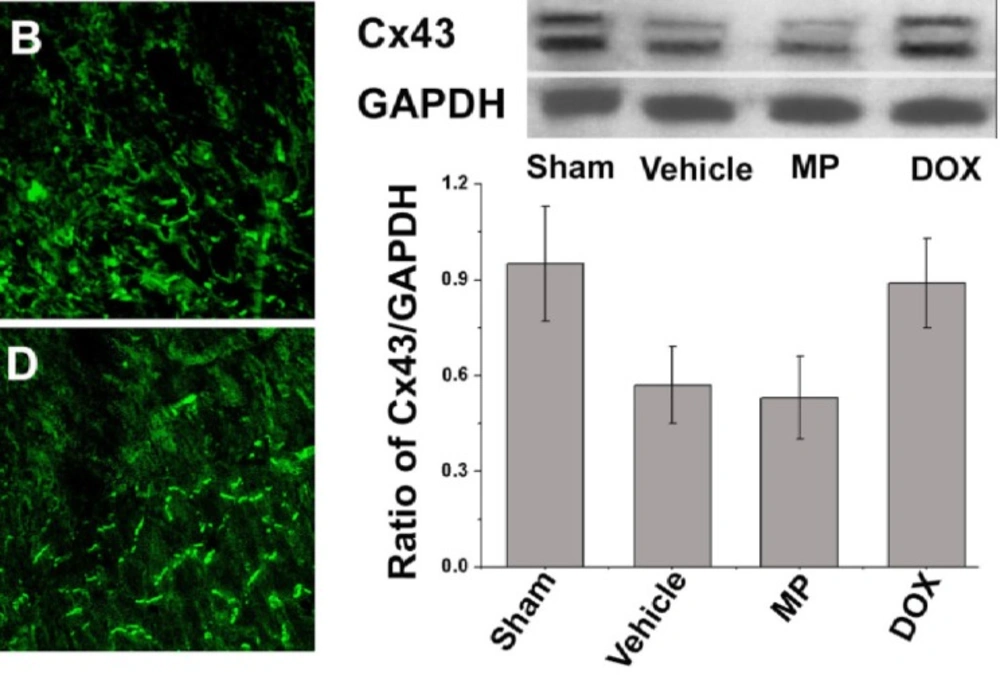
Immune fluorescent staining of Cx43
Immunofluorescence staining was conducted on the frozen section of the left ventricular central tissue sections of the infarcted rat hearts for 2 weeks using the method described in the literature (
15). Briefly, four specimens obtained from each group were marked with anti-Cx43 murine monoclonal antibodies (ab79010; Abcam, UK). Subsequently the sections were incubated with goat anti-rat IgG combined with FITC. Afterward, the sections were observed and photographed under confocal microscopy (Zeiss LSM510 META, Germany).
Western blot analysis of Cx43
Six heart samples were obtained from each group. The ventricular septa and right ventricle myocardia were cut into 1 mm sections along the grey region of the infarct and placed in liquid nitrogen for cryopreservation. The remaining myocardia were pulverized and boiled with SDS specimen buffer (0.25 M Tris–HCl, pH 6.8, 4% SDS, 40% glycerol, and 0.002% bromophenol blue) for 10 min after centrifugation. The proteins were then extracted. The equivalent protein was then purified via SDS-PAGE using 10% gel, and transferred on a nitrocellulose membrane. The membrane was blocked with 5% skimmed milk at room temperature for 2 h. Then, the proteins were incubated overnight with rabbit-derived anti-Cx43 (1:4000) and rat-derived anti-GAPDH (1:4000) at 4 °C. The samples were then incubated for 1 h with goat anti-mouse or anti-rabbit–HRP at room temperature. GADPH was used as the loading control. The density of each band was determined using the corresponding GAPDH value.
Data and statistics
All numerical data are presented as mean ± S.D. The number of animals in each group was not less than 18. The differences in echocardiographic measurements, western blot analysis, and MMP zymography were compared using one-way ANOVA followed by Turdey post hoc test. The occurrence of ventricular tachycardia was compared using a chi square test. The differences with p < 0.05 were considered significant.