Toll-like receptors (
TLRs) are the most important receptors in innate immunity that have been identified as a major class of pattern-recognition receptors. The
TLR family comprises at least eleven members; these
TLRs recognize a limited but highly conserved set of molecular structures (
1,
2).
TLR4, considered one of the most important TLRs, recognizes lipopolysaccharide (LPS) of Gram-negative bacteria. In addition to the LPS
, TLR4 recognizes heparan sulfate, heat shock proteins, and High Mobility Group Box1 (HMGB1) among other endogenous substances (
3). It has been reported that the ability of certain individuals to respond properly to
TLR4 ligands may be impaired by single-nucleotide polymorphisms (SNPs) within
TLR4 genes (
4-
6). Two cosegregating single nucleotide polymorphisms (SNPs) of the human
TLR4 gene, namely Asp299Gly (rs4986790) and Thr399Ile (rs4986791), have been correlated with a hyporesponsiveness to inhaled LPS (
7). The identification of
TLR4 Asp299Gly and Thr399Ile mutations might be important for the individual risk assessment of cancer patients treated by chemotherapy (
8,
9). Human colon cancers show heterogeneous behavior at the molecular and cellular levels, and the tumor cells can develop resistance to chemotherapy. 5-Fluorouracil (5-FU) is widely used for the treatment of patients with advanced colon cancers and is still the mainstay of chemotherapy (
10). However, this anticancer effect varies widely among individual patients. Some patients suffer adverse effects of 5-FU chemotherapy, while others do not. Therefore, the identification of the novel biological markers of 5-FU-resistant in tumor cells is one of the most important steps of developing optimal treatment strategies (
11,
12). Microbes or microbial products possess potent anticancer action (
13,
14). LPS, “a major component of the outer membrane of Gram-negative bacteria” is among the most potent TLR4 agonists which may affect the response to 5-FU chemotherapy in colorectal carcinoma. The Asp299Gly polymorphism with an allelic frequency of approximately 6% in individuals of mixed European descent (
1,
7) was found to prevent the binding of HMGB1 to
TLR4 in a dominant negative fashion (
15). The genotoxic agents that activate the Poly (ADP-ribose) polymerase (PARP), including alkylating agents, can stimulate the release of HMGB1 from its association with chromatin, most likely as a result of the direct PARP-mediated poly adenosyl ribosylation of HMGB1 (
16-
18). Chemotherapy-induced cell death triggers the release of the high-mobility group box 1 protein (HMGB1), which stimulates
TLR4 and elicits an immune response that is required for the success of the therapy (
4). One of the major goals of oncology is to predict the response of patients with cancer to chemotherapeutic agents by employing laboratory methods variously called ‹tumor chemosensitivity assays›, ‹drug response assays› or ‹drug sensitivity assays›
in-vitro (
19,
20). The present study evaluated the impact of two common single nucleotide polymorphisms (SNPs) of the human
TLR4 gene on the chemosensitivity of colon cancer cells to 5-fluorouracil.
A number of studies have revealed that HCT116 ‘colorectal carcinoma cell line’ do not express detectable amount of TLR4 (
21,
22). Therefore, HCT116 cells were chosen for transfection to eliminate the basal effects of TLR4 of the cells. HCT116 (ATCC, CCL-247) was obtained from ATCC and maintained in RPMI medium supplemented with 10% fetal bovine serum and 0.6% Pen-Strep at 37°C in a 5% CO
2 atmosphere (All cell culture reagents were purchased from Invitrogen, Carlsbad, CA, USA). LPS (
Escherichia coli 0111:B4) was purchased from Sigma-Aldrich and 5-FU was from Choongwae Pharma Corporation (Seoul, Korea). The levels of HMGB1 in the culture medium or cell lysate were analyzed by Western Blotting using rabbit monoclonal Ab (Abcam). Antihuman
TLR4 antibody against the surface
TLR4 was obtained from eBioscience. The apoptosis sampler kit containing rabbit-derived polyclonal antibodies (Abs) against caspase-3, PARP and the corresponding cleaved forms were obtained from Cell Signaling Technology. Apoptosis was determined using a commercial Annexin V-FITC apoptosis detection kit (BD Biosciences). Flag-CMV1-TLR4 wild-type and Flag-CMV1-TLR4 mutants (D299G and T399I) expression vectors were kindly provided as a gift by Prof. Dr. Vogel and Dr. Rallabhandi (University of Maryland). The DNA plasmid pmaxGFP (from Amaxa) was used as a positive control for transfection.
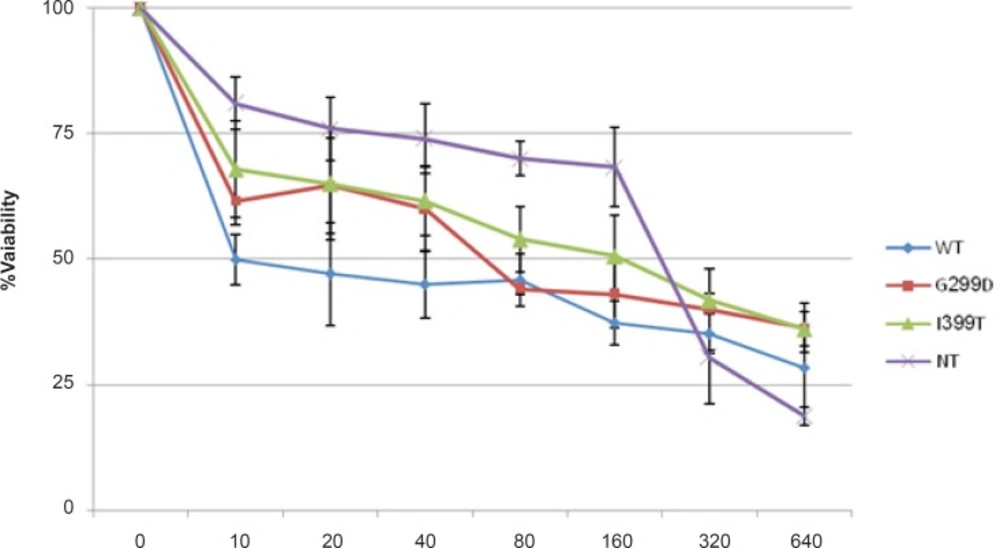
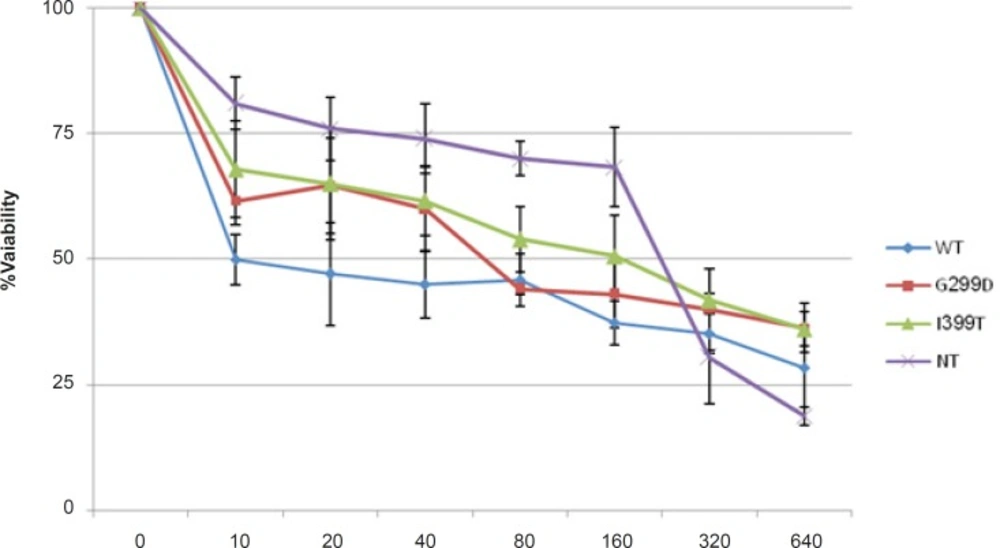
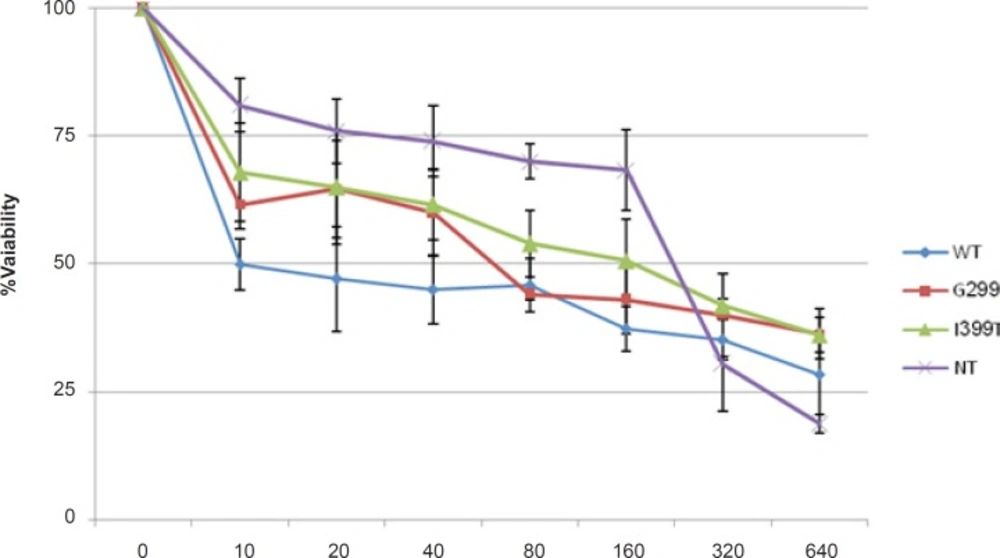
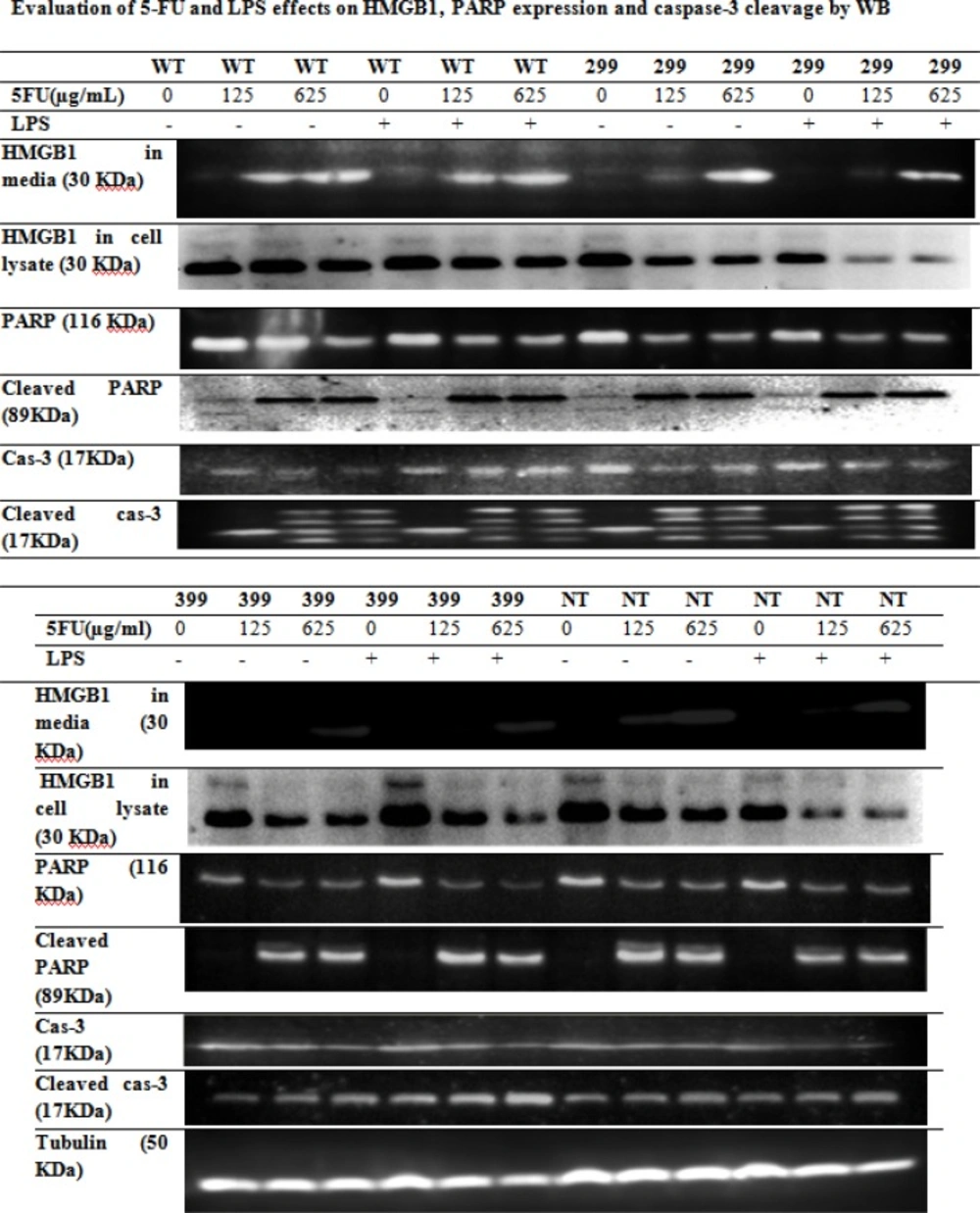
Transfected cells were treated with different concentrations of 5-FU for 48 h in the presence and absence of 1 μg/mL LPS. The culture supernatant was removed and frozen at - 80°C for detection of secreted HMGB1. Secreted HMGB1 in culture supernatant was concentrated by acetone (1.8 mL/200 μL supernatant) (
23). The level of HMGB1 in the culture medium and Whole-cell lysates were analyzed by western blot (WB) using rabbit monoclonal Ab (Abcam). Caspase-3 activity and Poly (ADP-ribose) polymerase (PARP) cleavage were also analyzed by WB (Rabbit, Cell signaling technology) in cell lysate.
Statistical analyses were conducted using the ANOVA general linear models procedure (GLM) of SAS software (SAS Institute, 2005). When ANOVA revealed significant effects, means were separated by Duncan’s multiple range tests. Data with p < 0.05 were considered to be significant.