
Introduction
Experimental
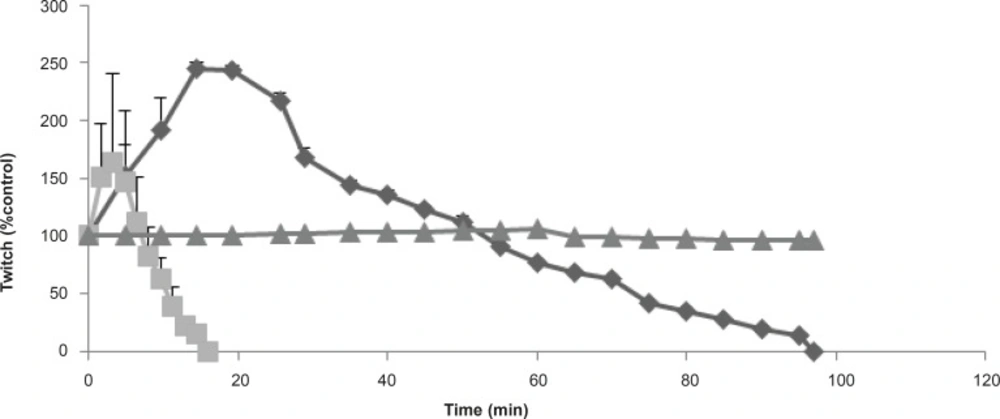
The effect of Buthotus schach scorpion venom at 1 μg/mL (▲), 3 μg/mL (♦); 10 μg/mL (■) concentrations, in response to indirect CBC stimulated preparations. Each point represents the maximum response for that concentration (mean ± SEM; n = 4).
Results and Descussion
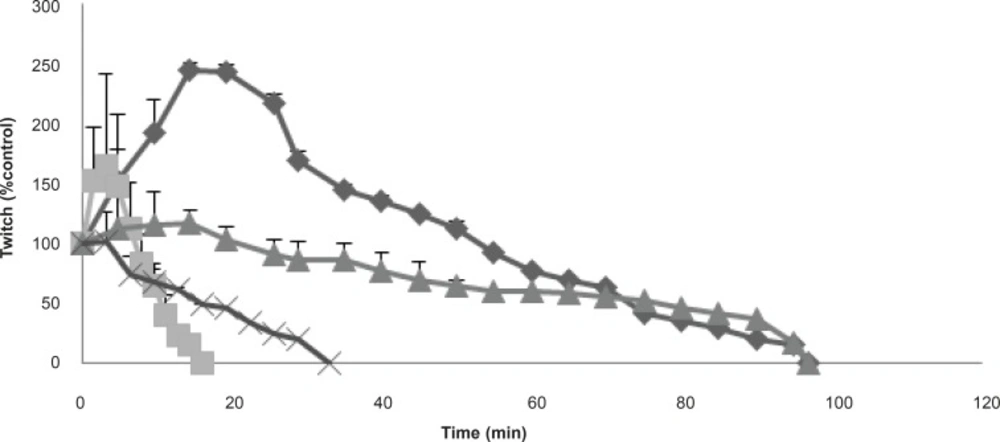
The effects of the venom at 3 μg/mL on twitch height in response to directly (▲) stimulated CBC preparations in compare with indirectly (♦) stimulated preparations are shown. Furthermore, its effects at 10 μg/mL are also shown on twitch height at both directly (●) and indirectly (■) CBC stimulated preparations. Each point represents the maximum response for that concentration (mean ± SEM; n = 4).

The effect of Buthotus schach scorpion venom at 3 μg/mL (■) and 10 μg/mL (□) concentrations in response to exogenous Ach (1-2 mM) for 30 sec, Carb (30-40 μM.) for 60 sec and KCl (20-40 mM) for 30 sec in CBC preparations. Each column represents the maximum response (% control) for that concentration. (Mean ± SEM; n = 4) (* = p < 0.05).
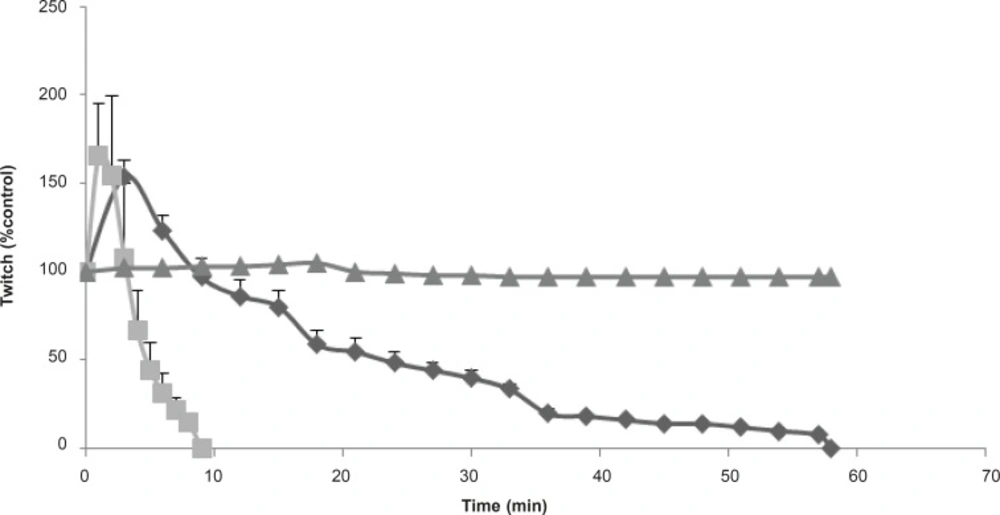
The effect of Buthotus schach scorpion venom at 1 μg/mL (▲), 3 μg/mL (♦); 10 μg/mL (■) concentrations, in response to indirect stimulation of MHD preparations. Each point represents the maximum response for that concentration (mean ± SEM; n = 4).
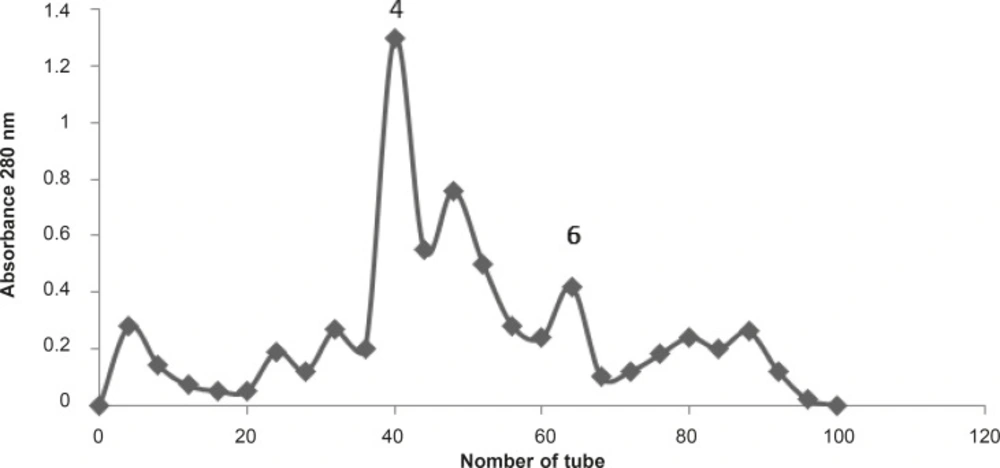
Buthotus schach scorpion venom (300 mg) was dissolved in Ammonium acetate buffer and loaded on Sephadex G50 column with 1 mL/min flow rate. Seven fractions were obtained. F4 and F6 are two pharmacologically active and responsible for the venom effects
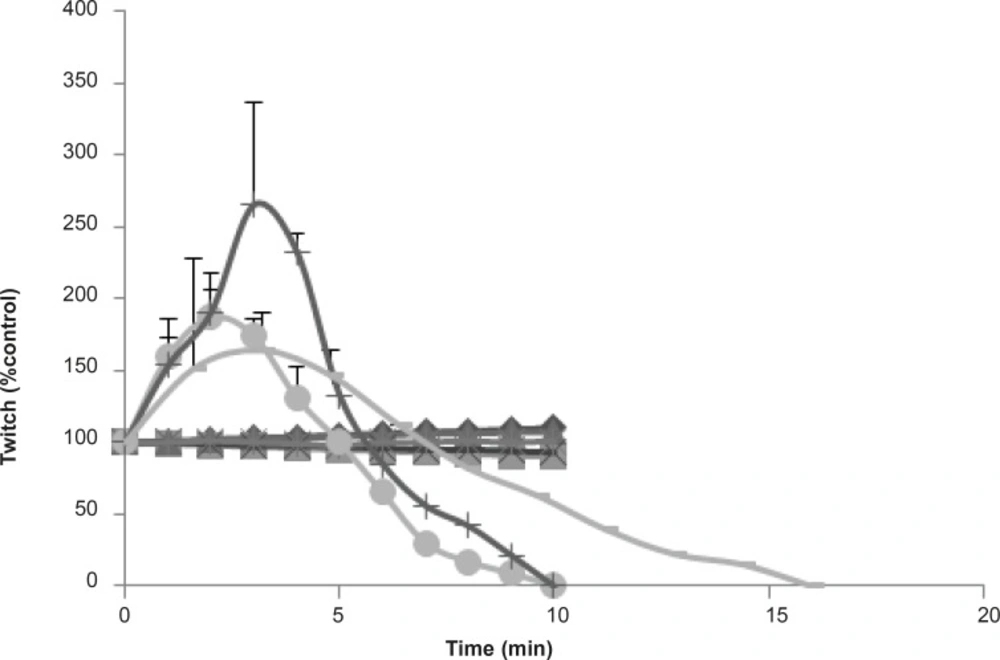
The effects of Buthotus schach scorpion venom and its seven collected fractions were examined each at 10 μg/mL concentrations on CBC indirectly stimulated nerve–muscle preparations. Fractions F1 (♦), F2 (■), F3 (▲), F5 (×), F7 (*), had shown no effects on twitch height or muscle contractions responses. However fractions F4 () and F6 (+) were shown a transient increase in twitch height followed by huge contracture led to muscle paralysis similar to one that was seen with the venom (●).
Discussion
| LD50 μg/mice | Max. Twitch height in response to indirect stimulation (% ctl) | Time to block NMJ (min) | |||||
|---|---|---|---|---|---|---|---|
| 1μg/mL | 3μg/mL | 10μg/mL | 1μg/mL | 3μg/mL | 10μg/mL | ||
| Crude venom | 84 | 106 | 244 | 164 | _ | 97 | 16 |
| F4 | 35/5 | 105 | 247 | 187 | _ | 20 | 10 |
| F6 | 21 | 105 | 217 | 265 | _ | 20 | 10 |
