
Introduction
Hypericin is one of the naphthodianthron constituents of St. John’s Wort (Hypericum perforatum L, SJW) belonging to the Hypericaceae family (1, 2). Clinical studies confirm that SJW extracts are effective for the treatment of mild to moderate depression and their effects are comparable to low dose tricyclic antidepressants (TCAs), but with less pronounced side effects (3). The antidepressant activity of SJW extracts has been attributed to hyperforin, hypericin and pseudohypericin and also to several flavonoids (3). From a pharmacological point of view, hypericin is the most interesting compound of H. perforatum L. (4, 5). It has been shown that the highest hypericin ratio was determined in flowers and buds generally collected between 8:00 and 10:00 AM within a day for examined hypericum populations (5). Butterweek et al. have suggested that adrenergic and/or serotonergic neurotransmission may be involved in antidepressant effect of TCAs and hypericin. Dopaminergic neurotransmission in the hypothalamus may also play an important role in mediating the antidepressant effects of SJW and hypericin (3). It has been shown that their antidepressant activity is related to inhibiting the reuptake of neurotransmitters such as serotonin, dopamine and norepinephrine (6). Hypericin also inhibits MAOA and MAOB activities in-vitro (7). St. John’s Wort can also cause serotonin syndrome when used in combination with the other drugs. This syndrome is the consequence of excess serotonergic activity in the central nervous system (8).
Conditioned Place Preference (CPP) is a widely-used experimental model for studying the rewarding properties of drugs in mice and rats. This animal model of drug reinforcement and drug dependence is suitable for studying the relationship between the rewarding stimulus properties of drugs and environmental stimuli. It has been demonstrated that the association of distinctive environmental stimuli with a primary reward such as food or a drug injection will result in an acquired preference for those specific environmental stimuli in the absence of the primary reward (9, 10). The majority of abused drugs, including morphine, readily condition a place preference in rodents. The production of rewarding and reinforcing effects is caused by the activation of μ-receptors; since reinforcement is antagonized by naloxone and the μ-receptor knock-out mice do not exhibit signs of morphine withdrawal (11). Disruption of morphine-induced conditioned place preference by naloxone suggests a role for opioid receptor mediation of the rewarding effects of morphine (9). The morphine-induced reinforcement effect could be due to the facilitation of dopaminergic transmission through stimulation of dopamine release (12-14). Release of dopamine from neurons in the presynaptic ventral tegmental area (VTA) into the nucleus accumbens causes reinforcement of the behavior (14). In addition to the dopaminergic system, opioidergic neurons interact with the serotonergic system. Serotonin is a potent stimulator of dopamine release (15, 16, 11). Therefore, an increase in brain serotonin could possibly stimulate the dopaminergic system (17). In this study, experiments were carried out to compare the effects of hypericin and synthetic antidepressants on the expression of morphine-induced conditioned place preference in rats.
Experimental
Animals
Adult male Wistar rats (Pasteur Institute, Tehran, Iran) used in the experiments, were 220-250 g at the beginning. Animals were housed four per cage and maintained at 24 ± 0.5°C with a controlled 12 h light-dark schedule with ad libitum food and water except during the experimental procedures. Each treatment group consisted of eight animals. There were seven or more days between the delivery of animals and the onset of experiments. Each animal was used only once and attention was paid to the ethical principles established in accordance with the committee of ethics of the Faculty of Medicine, Tehran University of Medical Sciences.
Methods
Animals were anesthetized through intra-peritoneal injections of xylazine (5 mg/Kg) and ketamine (80 mg/Kg) and placed into a stereotaxic device. An incision was made along the midline, the scalp retracted and the area surrounding bregma was cleaned and dried. In addition, lidocaine (2%) with epinephrine (0.3 mL) was injected into several locations around the incision for the purpose of local analgesia (19). A stainless steel guide cannula (22-gauge), 12 mm in length, was aimed at the left lateral ventricle (stereotaxic coordinates: 1.0 mm posterior to Bregma, 1.6 mm left lateral to midline, and 4.5 mm vertical from surface of the skull) (20). After cleaning the skull, a small amount of dental acrylic cement was pasted on to the surface so that it covered the skull screws and secured the implantation cannula in place. After the cement was completely dried and hardened, a stainless steel stylet was used to occlude the guide canula during the recovery and between drug injections. The incision was saturated and topical antibiotics were applied to the wound. The rat was removed from the stereotaxic apparatus and placed on a 37°C warming plate to allow them to recover from anesthesia. After surgery, the animals were individually housed and allowed to recover for seven days before experimental treatment. After the experiments were completed, cannula placement was confirmed by the infusion of 1% methylene blue solution (0.5 μL) and subsequent dissection. Figure 1 is confirming the canula placement.
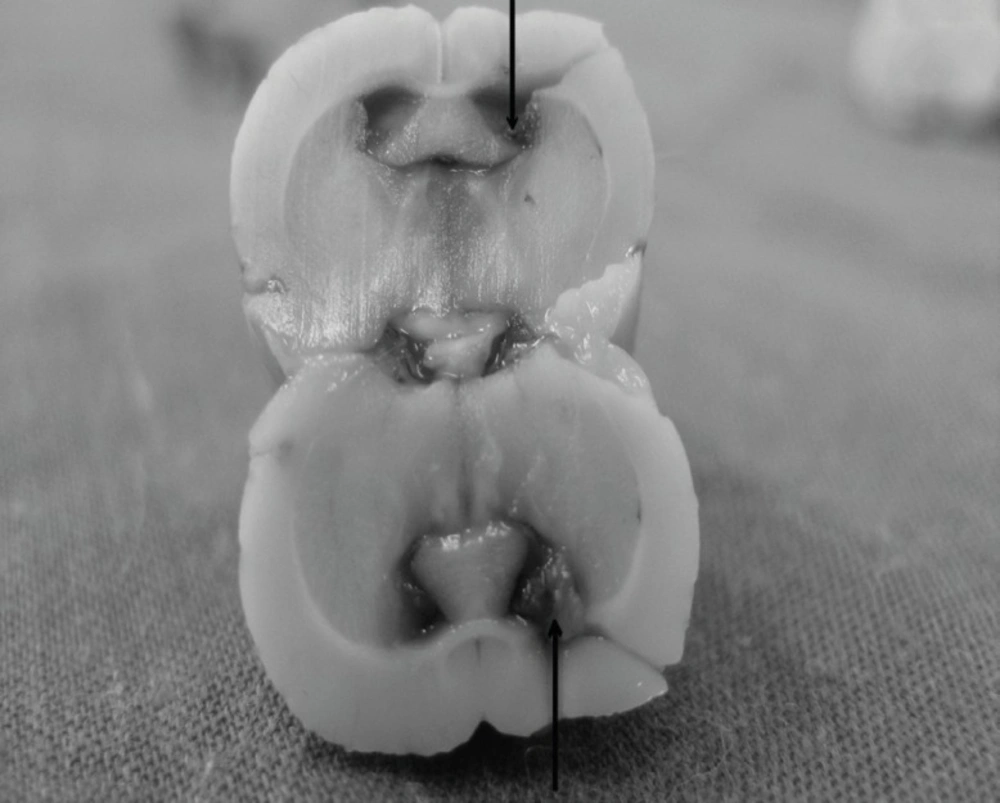
Canula placement confirmation by histological staining, the position has been pointed by a flash.
Materials
The used drugs were morphine sulphate (Temad, Iran), hypericin (Tocris, USA), fluoxetine hydrochloride (Cipla, India), imipramine hydrochloride (Sobhan Daru, Iran) and tranylcypromine sulphate (Goldshield Pharmaceuticals, South Africa). All drugs, with the exception of hypericin, were dissolved in saline; hypericin was dissolved in 1% ethanol and the solutions were injected IP or ICV. Hypericin is barely soluble in water; therefore, 5 mg of hypericin was nearly dissolved in 0.5 mL ethanol and further diluted with saline to a final concentration of 0.1 mg/mL. The ethanol concentration of the hypericin solution was 1% (21, 22). During the 2nd and 4th days, animals were injected with saline as a control for fluoxetine hydrochloride, imipramine hydrochloride and tranylcypromine sulphate.
Intracerebroventricular injection
Two μL of microsyringes were used to inject the drugs. Polyethylene tubing was used to attach the injection cannula to the microsyringe and 0.5 μL of hypericin, fluoxetine, imipramine or tranylcypromine solutions were delivered slowly over a 30 sec time period.
Apparatus procedure
A three-compartment place preference apparatus was made of Plexiglas, measuring 88 × 36 × 34 cm, consisting of two main compartments measuring 39 × 36 × 34 cm, one having grey sides with a smooth grey floor, the other having black and white stripes (2 cm wide) with a smooth white floor. The third compartment consisted of a white central platform measuring 10 × 36 × 34 cm, which was raised by 2 cm and separated the two main compartments. During the conditioning phase, compartments were isolated using guillotine doors (23).
Place conditioning
An unbiased CPP paradigm was used for six continuous days and consisted of three distinct phases including: preconditioning, conditioning and postconditioning. Animals were tested during the same time period (9:00 and 14:00 h) each day for each of the CPP paradigm phases (11, 24).
Preconditioning phase
On the first day of the trial, each rat was placed separately in the apparatus for 10 min with free access to all compartments and the time spent in each compartment was recorded to determine the least preferred side for the animals (11, 25, 19).
Conditioning phase
This phase involved four days. On the first and third days, animals were injected with morphine sulphate (5 mg/Kg) and confined to their least preferred compartment (white side) for 30 min. During the second and fourth days, animals were injected with saline and confined to their preferred compartment (grey side) for 30 min (14, 23).
Post-conditioning phase
This phase was carried out on the 6th day of the trials. Drugs (hypericin, fluoxetine, imipramine and tranylcypromine) were injected IP 15 min before or ICV 1 min before the beginning of this phase of the test (24, 11) and rats were allowed free access to all compartments for 10 min. No morphine injection was given during this phase (drug-free state). The time spent in the least preferred side (drug side) was recorded for each animal and the change in preference (CIP) was calculated as the difference (in sec) between the time spent in the drug side compartment during the post- and preconditioning phases (11, 14).
Measurement of conditioned place preference produced by morphine
Different doses of morphine were injected IP for the assessment of dose dependency of morphine-induced CPP.
Induction and assessment of place conditioning by morphine sulphate
In this experiment, the effect of morphine sulphate (2.5, 5 and 10 mg/Kg, IP) on producing place preference was tested. On the first and third days of the conditioning phase, animals received morphine and were placed in the drug side of the apparatus for 30 min. On the second and fourth days of the conditioning phase, animals received saline (1 mL/Kg, IP) and were placed in the preferred side of the apparatus for 30 min (14). It has been shown that CPP produced by morphine is dose-dependent and the submaximal and maximal responses are obtained by using 5 and 10 mg/Kg of morphine respectively (14, 24).
Measurement of the effect of hypericin on the expression of CPP induced by morphine sulphate
In order to test the effect of hypericin on the expression of morphine-induced CPP, this drug was injected IP (15 min) or ICV (1 min) before the postconditioning phase.
Measurement of the effect of synthetic antidepressant on the expression of CPP induced by morphine sulphate
In order to test the effect of fluoxetine, imipramine and tranylcypromine on the expression of morphine-induced CPP, these drugs were injected IP (15 min) or ICV (1 min) before the post-conditioning phase.
Statistical analyses
Values were reported as the mean change in preference ± SEM, and difference in time (sec) spent in the least preferred compartment before and after conditioning. One-way ANOVA followed by a Tukey test was used to calculate significance levels between the drugs. A value of p < 0.05 was considered significant.
Results and Discussion
Dose-response curve for CPP induced by morphine
Intra-peritoneal injection of morphine sulfate (2.5, 5 and 10 mg/Kg) in rats caused a dose-dependent CPP (Table 1). We used submaximal dose (5 mg/Kg) of morphine in this experiment. The saline control group showed no preference for either of the compartments (p < 0.0001).
| Phases | Saline (1 mL/Kg) | Morphine (2.5 mg/Kg) | Morphine (5 mg/Kg) | Morphine (10 mg/Kg) |
| Preconditioning | 209.30 ± 6.74 | 208.46 ± 2.26 | 201.77 ± 5.15 | 201.51 ± 4.60 |
| Postconditioning | 211.88 ± 5.84 | 224.05 ± 3.15 | 227.88 ± 5.19 | 238.44 ± 3.87 |
| Change in preference | 2.58 ± 1.07 | 15.59 ± 1.60*** | 26.10 ± 1.69*** | 36.92 ± 1.87*** |
Effects of IP injections of hypericin on the expression of morphine-induced CPP
As shown in Table 2, hypericin (25, 50 and 100 μg/Kg) increased the expression of morphine-induced (5 mg/Kg) CPP dose-dependently. The Saline and Saline + ethanol (1%) control group showed no preference for either of the compartments (p < 0.0001).
| Phases | Control | Morphine (5 mg/Kg) | |||
|---|---|---|---|---|---|
| Saline | Saline + Ethanol (1%) (1 mL/Kg) | Hypericin | Hypericin | Hypericin | |
| Preconditioning | 206.88 ± 2.47 | 208.70 ± 3.55 | 206.26 ± 3.30 | 203.42 ± 5.89 | 209.94 ± 4.72 |
| Postconditioning | 208.26 ± 2.31 | 211.98 ± 2.48 | 228.07 ± 2.49 | 235.02 ± 5.96 | 252.22 ± 5.69 |
| Change in preference | 1.38 ± 1.02 | 3.28 ± 1.28 | 20.56 ± 1.53*** | 31.60 ± 2.18*** | 42.27 ± 2.87*** |
Effects of IP injections of fluoxetine on the expression of morphine-induced CPP
Fluoxetine (2.5, 5 and 10 mg/Kg) increased the expression of morphine-induced (5 mg/Kg) CPP dose-dependently. The saline control group showed no preference (p < 0.0001) for either of the compartments (Table 3).
| Phases | Control | Morphine (5 mg/Kg) | ||
|---|---|---|---|---|
| Saline (1 mL/Kg) | Fluoxetine (2.5 mg/Kg) | Fluoxetine (5 mg/Kg) | Fluoxetine (10 mg/Kg) | |
| Preconditioning | 201.16 ± 11.25 | 198.53 ± 6.68 | 187.96 ± 13.24 | 193.44 ± 14.42 |
| Postconditioning | 203.22 ± 10.50 | 219.72 ± 5.49 | 221.94 ± 13.31 | 238.09 ± 12.80 |
| Change in preference | 2.06 ± 1.18 | 21.18 ± 1.61*** | 33.98 ± 1.19*** | 44.64 ± 2.81*** |
Effects of IP injections of imipramine on the expression of morphine-induced CPP
Imipramine (5, 10 and 20 mg/Kg) increased the expression of morphine-induced (5 mg/Kg) CPP dose-dependently. The saline control group showed no preference (p < 0.0001) for either of the compartments (Table 4).
| Phases | Control | Morphine (5 mg/Kg) | ||
|---|---|---|---|---|
| Saline (1 mL/Kg) | Imipramine 5 mg/Kg | Imipramine 10 mg/Kg | Imipramine 20 mg/Kg | |
| Preconditioning | 203.61 ± 5.01 | 221.89 ± 6.86 | 206.47 ± 11.37 | 203.63 ± 5.46 |
| Postconditioning | 207.50 ± 4.35 | 241.77 ± 6.74 | 237.31 ± 11.90 | 244.08 ± 4.29 |
| Change in preference | 3.89 ± 1.56 | 19.88 ± 0.66*** | 30.84 ± 1.84*** | 40.44 ± 1.46*** |
Effects of IP injections of tranylcypromine on the expression of morphine-induced CPP
Tranylcypromine (5, 10 and 20 mg/Kg) increased the expression of morphine-induced (5 mg/Kg) CPP dose-dependently. The saline control group showed no preference (p < 0.0001) for either of the compartments (Table 5).
| Phases | Control | Morphine (5 mg/Kg) | ||
|---|---|---|---|---|
| Saline | Tranylcypromine | Tranylcypromine | Tranylcypromine | |
| Preconditioning | 206.02 ± 5.95 | 209.73 ± 8.43 | 207.54 ± 4.93 | 214.53 ± 5.24 |
| Postconditioning | 209.05 ± 5.42 | 234.66 ± 7.72 | 243.34 ± 4.54 | 265.52 ± 6.36 |
| Change in preference | 3.03 ± 0.90 | 24.92 ± 2.70*** | 35.80 ± 1.94*** | 50.98 ± 1.80*** |
Effects of ICV injections of hypericin on the expression of morphine-induced CPP
ICV injections of hypericin (0.025, 0.05 and 0.1 μg/rat) increased the expression of morphine-induced (5 mg/Kg) CPP. The saline and saline + ethanol (1%) control group showed no preference (p < 0.0001) for either of the compartments (Table 6).
| Phases | Control | Morphine (5 mg/Kg) | |||
|---|---|---|---|---|---|
| Saline | Saline + Ethanol 1% | Hypericin | Hypericin | Hypericin | |
| Preconditioning | 215.51 ± 3.53 | 216.11 ± 3.14 | 201.94 ± 4.79 | 191.62 ± 2.68 | 195.97 ± 5.37 |
| Postconditioning | 215.51 ± 3.47 | 220.64 ± 3.61 | 248.33 ± 5.62 | 265.61 ± 6.17 | 294.78 ± 4.49 |
| Change in preference | 1.23 ± 0.88 | 4.53 ± 1.57 | 46.39 ± 3.72*** | 73.99 ± 4.90*** | 96.46 ± 5.34*** |
Effects of ICV injections of fluoxetine on the expression of morphine-induced CPP
ICV injections of fluoxetine (2, 5 and 10 μg/rat) increased the expression of morphine-induced (5 mg/Kg) CPP. The saline control group showed no preference (p < 0.0001) for either of the compartments (Table 7).
| Phases | Control | Morphine (5 mg/Kg) | ||
|---|---|---|---|---|
| Saline (1 mL/Kg) | Fluoxetine (2 μg/rat) | Fluoxetine (5 μg/rat) | Fluoxetine (10 μg/rat) | |
| Preconditioning | 204.03 ± 11.66 | 196.59 ± 6.33 | 181.13 ± 7.86 | 190.28 ± 14.60 |
| Postconditioning | 203.72 ± 10.37 | 221.76 ± 5.06 | 251.71 ± 11.64 | 266.19 ± 11.63 |
| Change in preference | -0.31 ± 1.89 | 25.16 ± 4.73*** | 70.58 ± 6.89*** | 75.91 ± 3.52*** |
Effects of ICV injections of imipramine on the expression of morphine-induced CPP
ICV injections of imipramine (5, 10 and 20 μg/rat) increased the expression of morphine-induced (5 mg/Kg) CPP. The saline control group showed no preference (p < 0.0001) for either of the compartments (Table 8).
| Phases | Control | Morphine (5 mg/Kg) | ||
|---|---|---|---|---|
| Saline (1 mL/Kg) | Imipramine (5 μg/rat) | Imipramine (10 μg/rat) | Imipramine (20 μg/rat) | |
| Preconditioning | 208.32 ± 4.98 | 202.44 ± 8.45 | 207.94 ± 6.63 | 203.98 ± 6.09 |
| Postconditioning | 210.41 ± 5.50 | 235.54 ± 9.12 | 253.39 ± 6.64 | 264.09 ± 5.52 |
| Change in preference | 2.09 ± 1.03 | 33.09 ± 0.92*** | 45.45 ± 1.82*** | 60.11 ± 2.43*** |
Effects of ICV injections of tranylcypromine on the expression of morphine-induced CPP
ICV injections of tranylcypromine (5, 10 and 20 μg/rat) increased the expression of morphine-induced (5 mg/Kg) CPP. The saline control group showed no preference (p < 0.0001) for either of the compartments (Table 9).
| Phases | Control | Morphine (5 mg/Kg) | ||
|---|---|---|---|---|
| Saline (1 mL/Kg) | Tranylcypromine (5 μg/rat) | Tranylcypromine (10 μg/rat) | Tranylcypromine (20 μg/rat) | |
| Preconditioning | 211.93 ± 3.40 | 212.61 ± 6.69 | 199.45 ± 3.44 | 200.48 ± 5.96 |
| Postconditioning | 212.79 ± 3.55 | 247.61 ± 5.54 | 247.48 ± 13.22 | 275.21 ± 5.71 |
| Change in preference | 0.86 ± 1.02 | 35.00 ± 2.17*** | 48.02 ± 11.18*** | 77.61 ± 3.38*** |
In the present study, the effects of hypericin and synthetic antidepressants on the expression of morphine-induced CPP have been compared. A 2008 detailed review of 29 randomized, placebo controlled trials found that St. John’s Wort was consistently more effective than placebo and equally effective to standard antidepressants (26). It has been shown that hypericin in 0.4-2.7 mg/day given 4-6 weeks to patients with mild to moderate depression compared to other antidepressants have similar treatment responders (26). It has been suggested that hypericin inhibits the reuptake of serotonin, dopamine and norepinephrine, and is as effective as standard antidepressants but with fewer side effects (3, 6). Hypericin, like synthetic antidepressants, produces serotonin syndrome when used concurrently with other serotonin reuptake inhibitors. Recently, case reports of such events have begun to trickle in. This is a potentially serious risk and SJW should not combine with prescription antidepressants except on the specific advice of a physician (27, 28, 29). Morphine produces a reward effect, which could be due to the facilitation of dopaminergic transmission by stimulating the release of dopamine (30, 12). Serotonin is a potent stimulator of dopamine release (30, 16).
Conclusion
In conclusion, the results of this study show that IP and ICV injections of hypericin and synthetic antidepressant produced a morphine-like effect on CPP and this effect may be related to increasing dopamine and serotonine concentration in synaptic clefts
