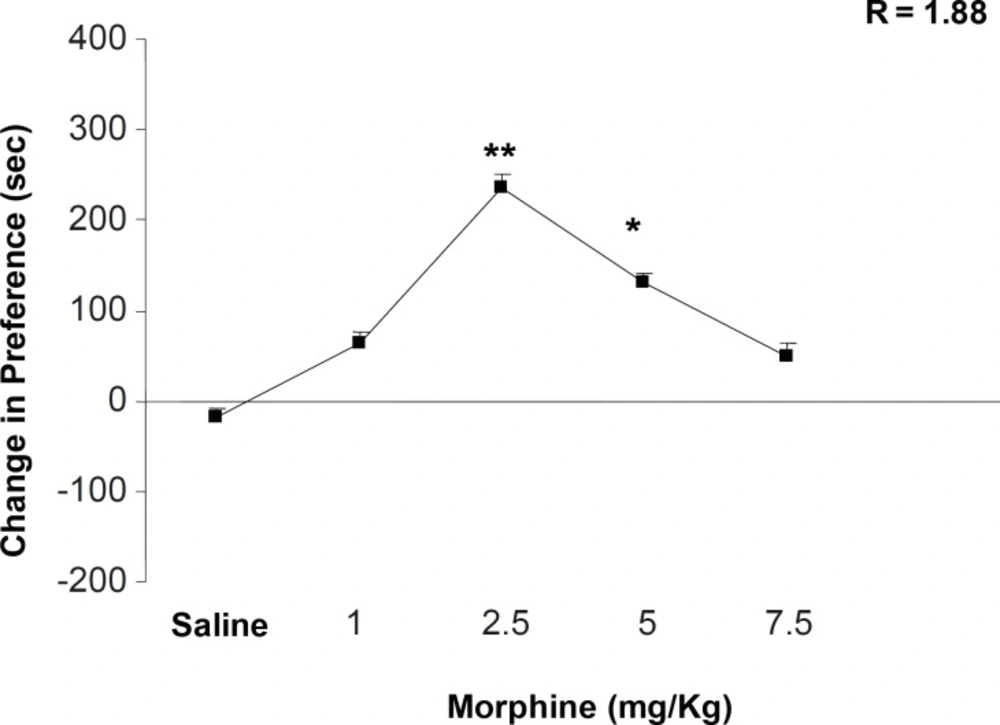
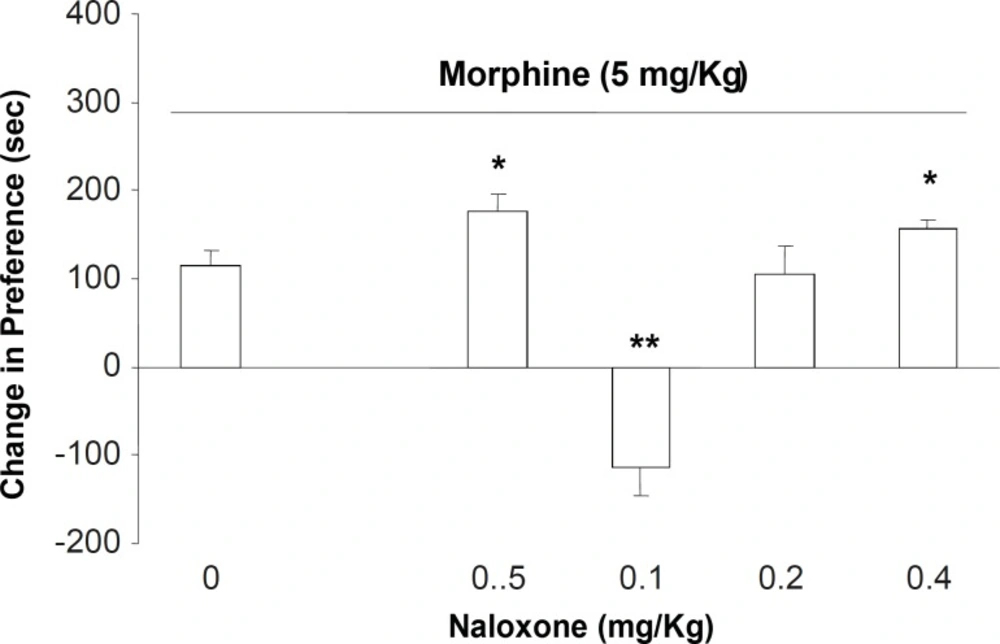
Our results indicated that the inhibition of opioid receptors by naloxone could potentiate the expression of morphine-induced place preference conditioning in doses of 0.05 and 0.4 and reverses the opioid effect in dose of 0.1 mg/Kg. It is important to notify that the similar results obtained for low and high doses of naloxone may not have similar base and mechanism(s). On the other hand, numbers of sniffing, rearing, and locomotion as indicators of mesolimbic dopamine-system activity (
15-
20) were not different form the control group. Results obtained from our experiments indicated that naloxone in low and high doses exacerbate and in medium dose (
e.g., 0.1 mg/Kg) reversed the expression of morphine CPP. These findings indicated the involvement of opioid receptors in the phenomenon. In agreement with previous studies, our results indicated that morphine can induce place preference in rats and mice (For rev see:
6,
21,
22). However, our results indicated that the maximum effect on place preference was achieved in dose of 2.5 mg/Kg which is also reported in previous studies (
6). Previous studies also indicated that opioid receptor inhibition reduced the acquisition and expression of place preference induced by opioids (
6,
23,
24,
25,
26,
27). On the other hand, in the previous studies, the effects of antagonists were observed on the doses of opioids which induced maximum response (
6,
24,
27); this response was achieved at doses in which probably just one type of the opioid receptors was occupied by the opioid (
6,
13,
24), though, in the present study, we proposed that by blockade of opioid receptors when downward shift of morphine dose-response curve was begun, dissociation the influence of different opioid receptors could be achieved on the morphine place preference. The special point here is that the receptor shifting may be occurring. In agreement with our proposal, studies have shown that agonists of kappa opioid receptors can induce place aversion in mice (
6,
23,
27). It would have been quite normal that if these receptors were inhibited (by naloxone for example), the effects of other opioid receptors could have been observed. As the rewarding effects of opioids are related to mu-opioid receptor activation (
23), in the present study, one can conclude that inhibition of the other opioid receptors is the main reason for augmentation of the expression of morphine-induced place preference which was observed for high naloxone (0.4 mg/Kg) dose. It seems that the reason of potentiating the expression of morphine-induced place preference by low dose of naloxone (
e.g. 0.05 mg/Kg) is different and needs another explanation. However, it is now clear that mu-opioid receptors can either inhibit or activate the enzyme adenylate cyclase (AC) in the plasma membrane of the target cells (
13,
28), which in turn, decreases or increases intra-cellular concentration of the cyclic adenosine monophosphate (cAMP), respectively (
13,
28). Although the population of the last kind of receptors is much lower, activation of this kind of opioid receptors can unfortunately mask the real potential of the seconds (
28). Therefore, inhibition of these receptors by low doses of opioid receptor antagonists can increase morphine antinociception as well as reward potency (
13,
29,
30). Our explanation for the effect of low dose of naloxone is that the drug may inhibit the mu-opioid receptors which activate AC and unmasks the real role of the main mu-opioid receptor population which inhibit AC (
13). So, the potentiation of the expression of morphine-induced place preference was observed as a result.
One important finding from our results is that the effects were not due to a change in dopamine-mesolimbic system activity as the behaviors related to the activities of mesolimbic dopamine system (dopamine related behaviors) such as sniffing, rearing and locomotion did not changed for the experimental and control groups. In addition, total compartment entering as an indicator of decision making was not changed which indicated that the behaviors related (at least in part) to the other central nervous system regions (i.e. frontal cortex) was not affected by our intervention.
In conclusion, our results indicated that the expression of morphine place preference may be related to the mu-opioid receptors activity. Besides, the receptors may also be similar to opioid receptors that inhibit AC rather than those activate the enzyme.