Animals and housing conditions:
Pregnant Albino Wistar rats (Pasteur institute, Tehran, Iran) were purchased and kept under standard laboratory conditions with free access to food and water at animal facility center of the Department of Exercise Physiology, Central Tehran Branch, Islamic Azad University. Neonatal maternal separation protocol was applied to animals based on previous studies (
17,
22). In brief, postnatal day 0 (PND 0) was considered as the day of birth and male pups were subjected to MS stress. The pups were briefly handled and then were separated from their mothers for 3 h every day during PND 2 - PND 14, beginning at 09:00 a.m. At the PND 21, male animals were housed in groups (four rats / cage) until the (PND 28). Institutional guidelines were strictly followed for animal care and use (Department of Exercise Physiology, Central Tehran Branch, Islamic Azad University, Tehran, Iran).
Materials and treatments:
Fluoxetine hydrochloride (Sigma USA) was dissolved in saline and solution was prepared instantly before the injections. Fluoxetine (FLX) was administered intraperitoneally (i.p.) in a constant volume of 5 ml/kg body weight to both MS and the control rats from PND 28 to PND 60. We used chronic administration of FLX (5 mg/kg) to MS and control rats because it has been reported as an effective treatment for modifying depressive-like behaviors in rodents (
17,
23).
Running wheel (RW) exercise:
At the PND 21, the animals were randomly subjected to exercise protocols including RW exercise. In case of RW exercise, the MS and control rats were allowed to adapt to the RW apparatus for 1 week. The exercise protocol was carried out based on the method previously described (
17,
24). In order to avoid the effects of social isolation, two rats in a cage were kept apart with a perforated Plexiglas separator between them that facilitated communication between the rats. The control group of MS rats were similarly handled but had no access to RW. Each activity wheel was made of Plexiglas (circumference = 105 cm, width = 10 cm, Novidan Tab, Iran) which was freely rotated against a resistance of 5g. Each wheel was connected to a magnetic switch that was linked to a counter placed outside of the animals’ cage and monitored the revolutions per hour. The rats had free access to the activity wheel 24 h a day for 32 days until PND 60, and daily running distance (km) was recorded.
Behavioral tests
Open-field test (OFT)
This test was used to evaluate the effects of housing and treatment conditions on number of squares crossed (horizontal activity) and the number of rearing (vertical activity) based on our previous studies (
8,
17).
Forced swimming test (FST)
The passive behavior (immobility time) was defined as the main behavioral marker in depression according to Porsolt
et al., 1978 (
25) in cylindrical glass tank (50×20 cm in diameter) containing 30 cm of water (23 ± 1 °C). Two swimming sessions were performed with an initial 15 min ‘pre-test’ followed by a 5 min ‘test’ after 24 h.
Splash test
The anhedonic state and self-care difficulties in animals in splash test as described by our previous studies based on recording the first grooming latency and the time spent grooming for a period of 5 min (
8,
17). A 10% sucrose solution was squirted on the dorsal coat of animals in their home cage. The first grooming latency and the time spent grooming were recorded for a period of 5 min.
Mitochondrial functional tests
Mitochondrial preparation
Different sets of animals from those used in the behavioral experiments were used to investigate the mitochondrial function and oxidative state in the HIPP. Hippocampi were quickly dissected out on ice after decapitation, soaked in the liquid nitrogen, and kept at -80 °C until the assays. Measurement of the protein concentration was performed based on Coomassie blue protein-binding method using bovine serum albumin (BSA) as the standard. The extraction of mitochondrial fractions was carried out as previously described (
26,
27). In order to confirm the purity of mitochondria, succinate dehydrogenase (SDH) activity was measured (
28).
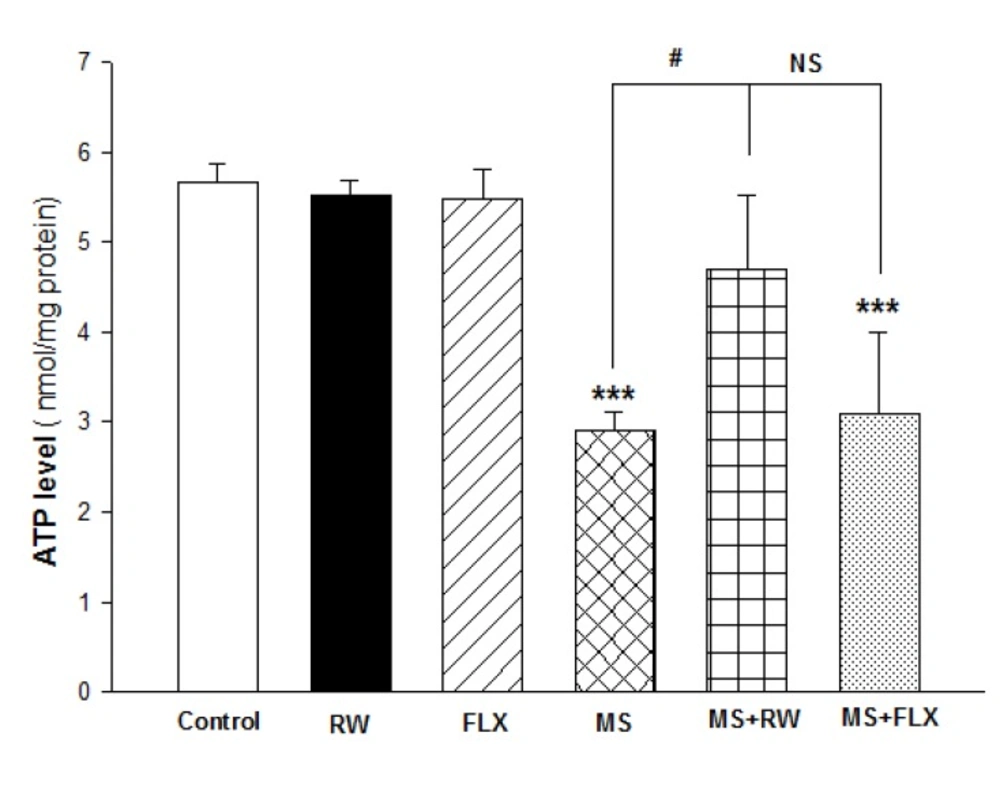
Adenosine triphosphate (ATP) assay
The ATP levels were measured using luciferase enzyme as described in our previous work (
29,
30). Bioluminescence intensity was measured using Sirius tube luminometer (Berthold Detection System, Germany).
Measurement of reactive oxygen spices (ROS) formation
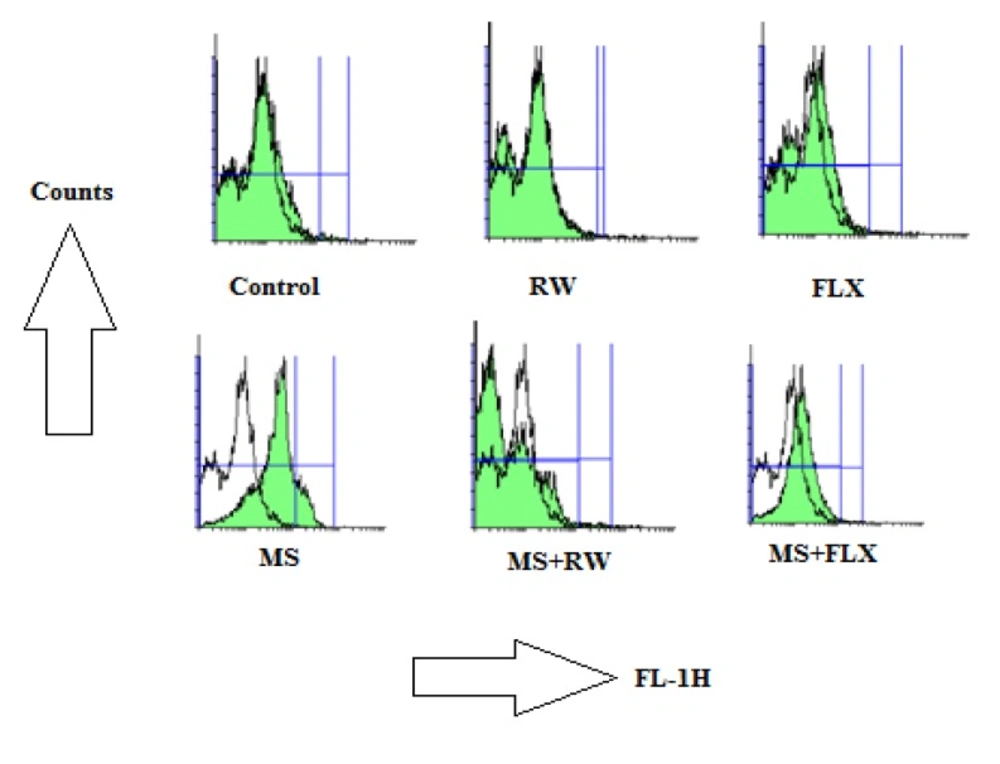
In order to measure the mitochondrial ROS formation, isolated HIPP mitochondria was incubated with DCFH-DA(10 µM) in respiratory buffer using the BD Biosciences FACS Caliber TM flowcytometer using a 530-nm band pass filter (FL-1 channel) (
31,
32).
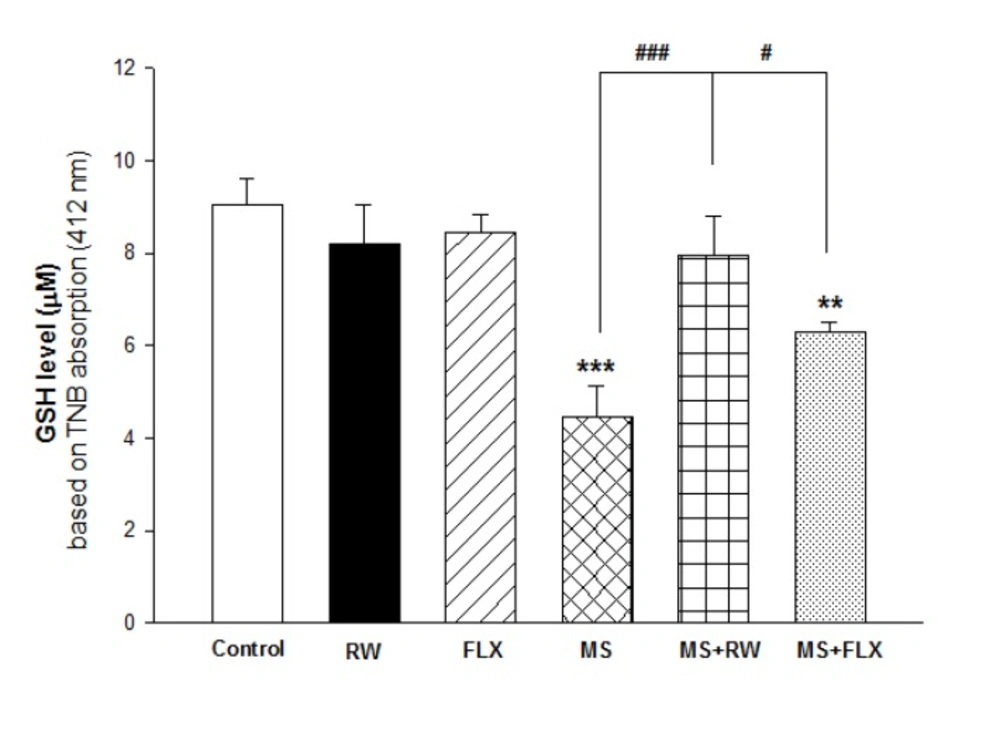
Reduced glutathione (GSH) level assay:
Glutathione levels were determined using of 5, 5′-dithiobis- (2-nitrobenzoic acid) or 5′-dithiobis-2-nitrobenzoic acid (DTNB)as a reagent with developed yellow color at 412 nm using a spectrophotometer (UV-1601 PC, Shimadzu, Japan) expressed as µM/mg protein based on the calibration standard curve (
32,
33).
Experimental design:
In this study, the animals were divided into six experimental groups each contains 6-8 male rats as follows; 1) Control group, 2) MS group, 3) Control group + RW treatment, 4) Control group + FLX treatment 5) MS+FLX treatment, and, 6) MS+RW treatment. After the termination of exercise protocol and FLX treatment at PND 60, the animals were subjected to behavioral tests. Also, at PND 60, the animals in the experimental groups were sacrificed under mild anesthesia and HIPP tissues were freshly collected for mitochondrial activity assays as described above.
Statistics
Comparison between the groups was analyzed using t-test and one-way ANOVA followed by multiple comparison tests in the Graph-pad prism software (version 6). P ˂ 0.05 was considered statistically significant.