
Introduction
Experimental
Results
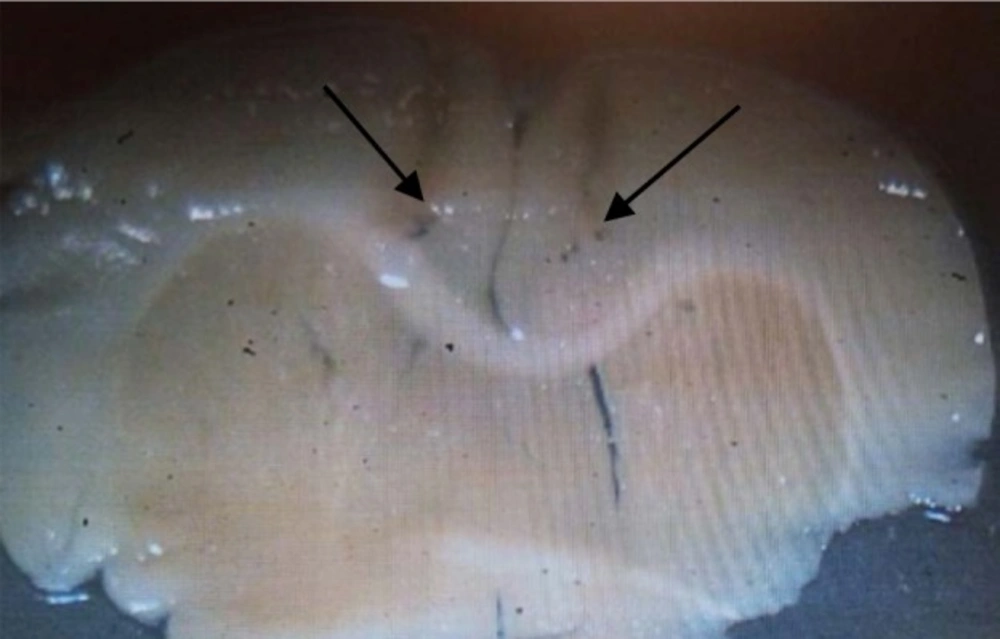
micrograph image of the injection site in the PL region
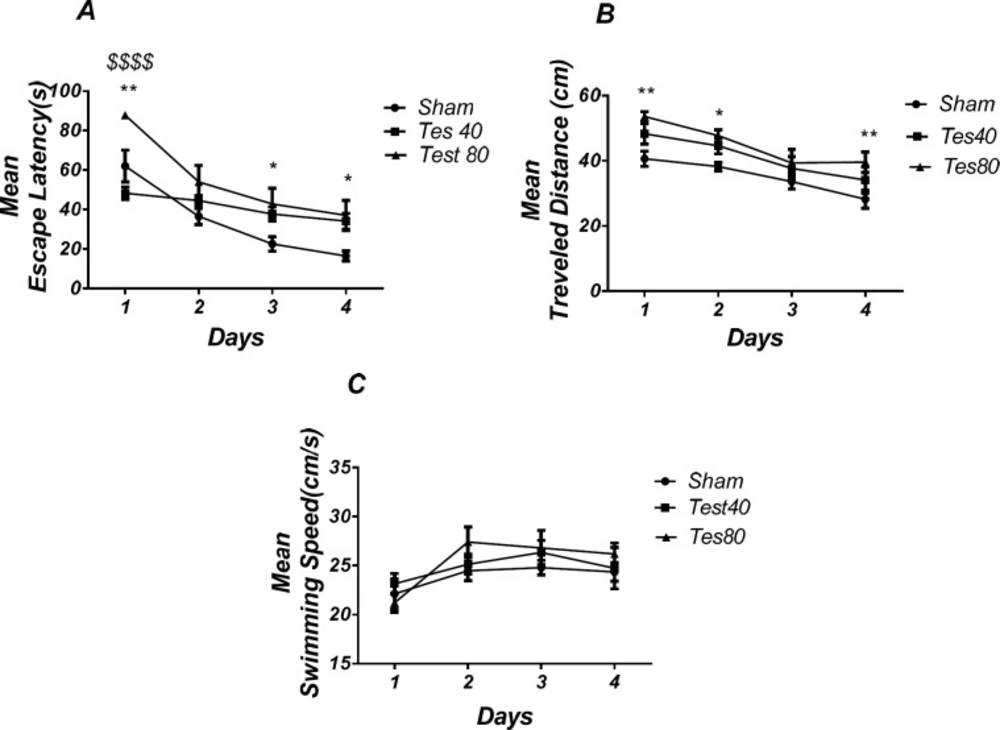
Effect of testosterone on spatial learning and memory. There was a significant increase in escape latency in1st (** P < 0.01), 3rd and 4th (* P < 0.05) days in testosterone treated animals compared to the sham operated group. Testosterone 80 treated animals showed a significant increase in 1st ($$$$ P < 0.001) day compared to the testosterone 40 treated animals group (A). Also, there was a significant increase in traveled distance in1st and 4th (* P < 0.05) and 2nd (* P < 0.05) days in testosterone treated animals compared to the sham operated group (B). No significant difference was found in swimming speed among different groups (C). (RM) Two-way analysis of variance (ANOVA) followed by post hoc analysis (Tukey test) were used and P < 0.05 was considered to be statistically significant. (n = 6-8 for each group)
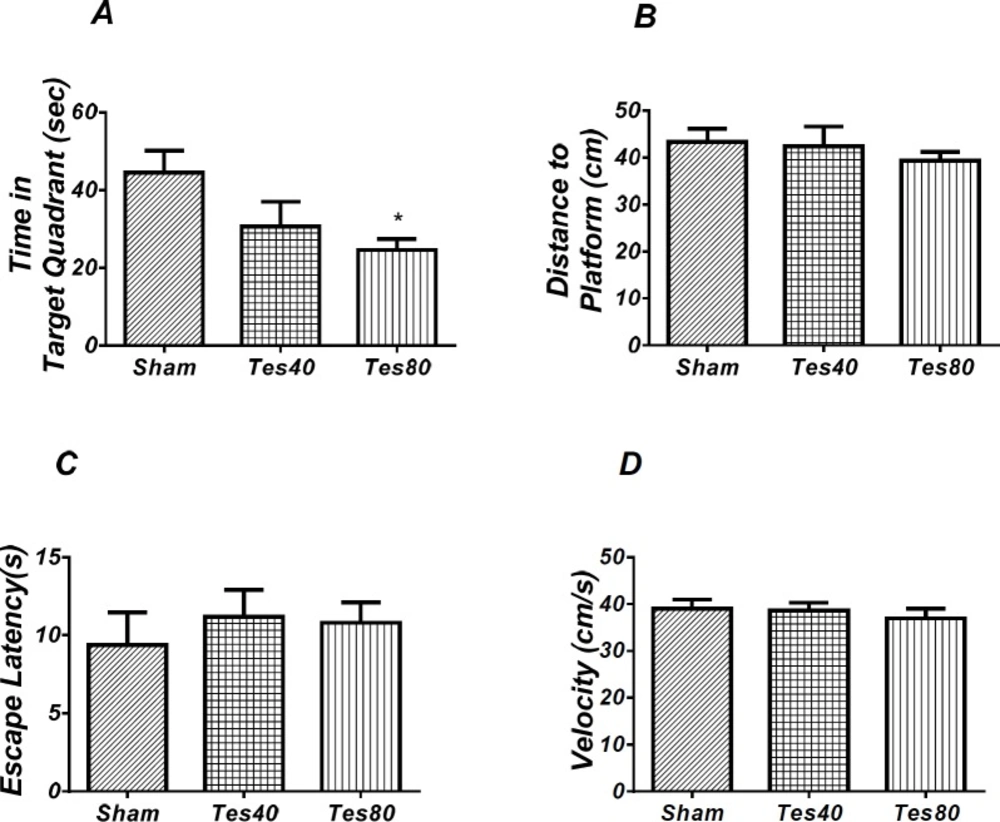
Effect of testosterone on probe test spatial learning and memory and visible test. There was significant decrease in time spent in the target quadrant in animal received testosterone 80 compared to the sham operated group (* P < 0.05) (A). There was no significant difference in distance to platform (B). In visible platform test there were no significant differences in escape latency (C) and velocity (D). One-way analysis of variance (ANOVA) followed by post hoc analysis (Tukey test) were used and P < 0.05 was considered to be statistically significant. (n = 6-8 for each group)
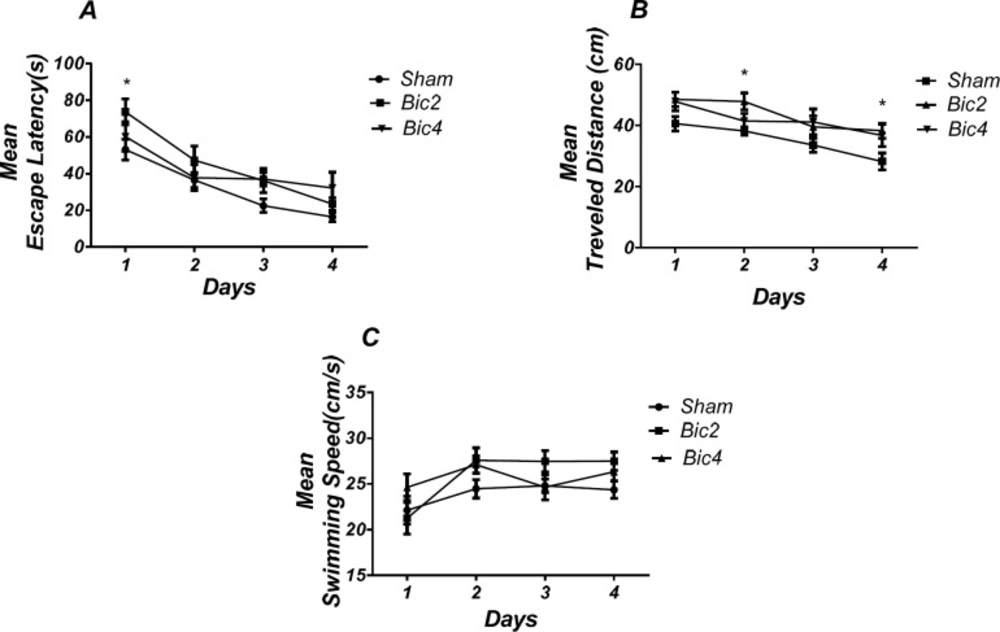
Effect of bicuculline on spatial learning and memory. There was a significant increase in escape latency in1st (* P < 0.05) day in bicuculline treated animals compared to the sham operated group (A). Also there was a significant increase in traveled distance in 2nd and 4th (* P < 0.05) days in bicuculline treated animals compared to the sham operated group. (B) No significant difference was observed in swimming speed among different groups (C). (RM) Two-way analysis of variance (ANOVA) followed by post hoc analysis (Tukey test) were used and P < 0.05 was considered to be statistically significant. (n = 6-8 for each group)
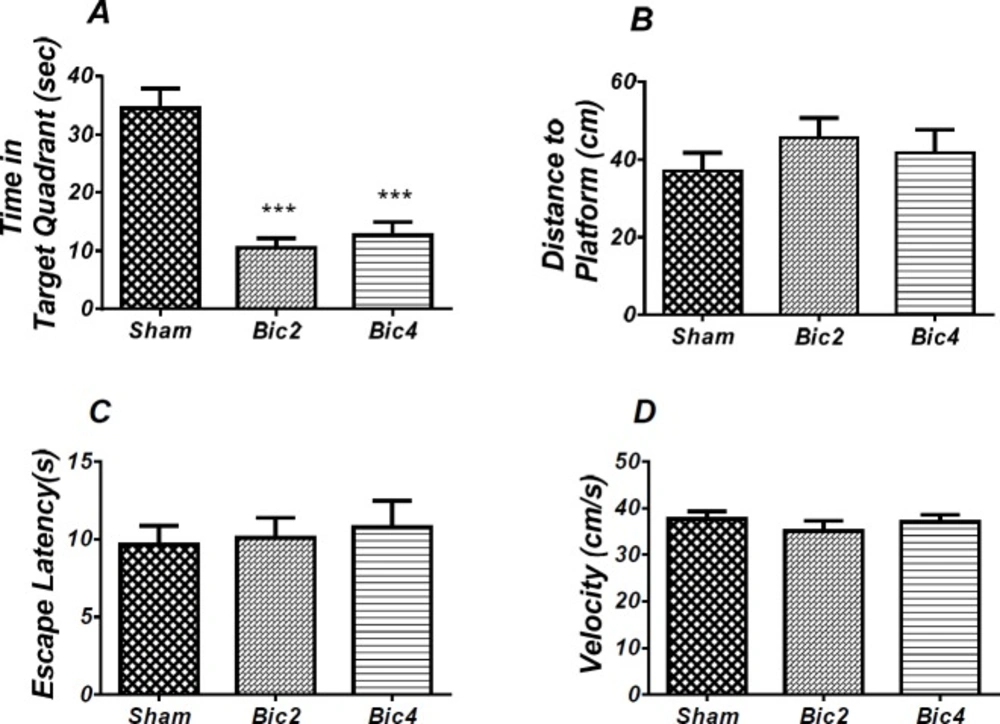
Effect of bicuculline on probe test spatial learning and memory and visible test. There was significant decrease in time spent in the target quadrant in animal received bicuculline compared to the sham operated group (*** P<0.001) (A). There was no significant difference in distance to platform (B). In visible platform test there were no significant differences in escape latency (C) and velocity (D). One-way analysis of variance (ANOVA) followed by post hoc analysis (Tukey test) were used and p < 0.05 was considered to be statistically significant. (n = 6-8 for each group)
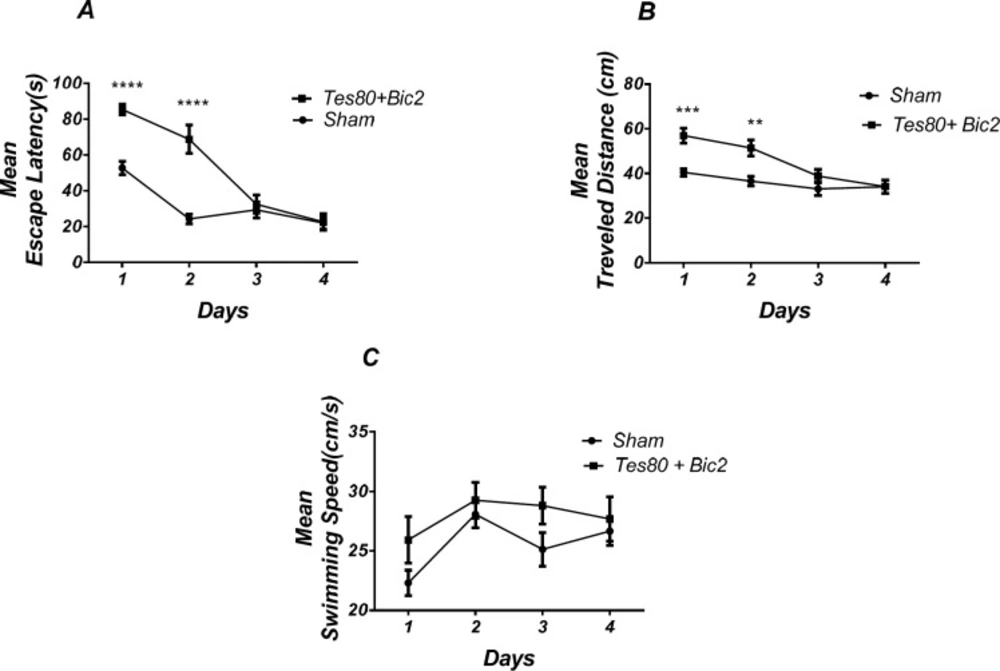
Effect of testosterone + bicuculline on spatial learning and memory. There was a significant increase in escape latency in1st and 2 nd (*** P < 0.001) days in testosterone + bicuculline treated animals compared to the sham operated group (A). Also, there was a significant increase in traveled distance in 1st (*** P < 0.001) and 2 nd (** P < 0.01) days in testosterone + bicuculline treated animals compared to the sham operated group (B). No significant difference was observed in swimming speed among groups (C). (RM) Two-way analysis of variance (ANOVA) followed by post hoc analysis (Bonferroni’s test) were used and P < 0.05 was considered to be statistically significant. (n = 6-8 for each group)
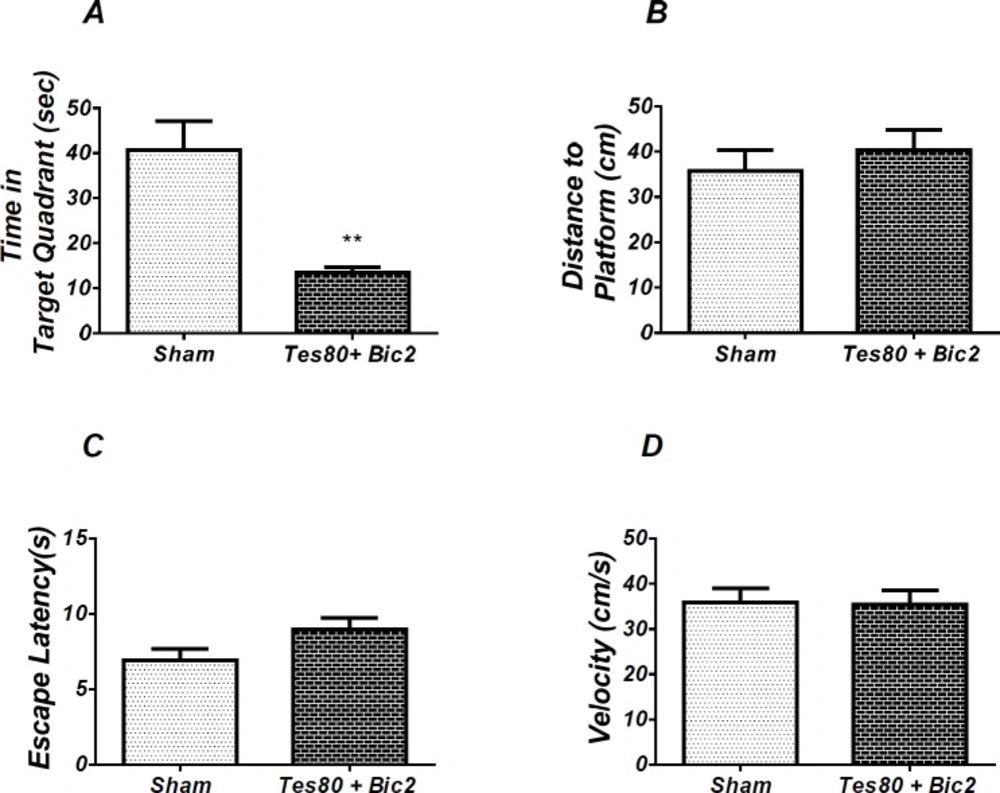
Effect of testosterone + bicuculline on probe test spatial learning and memory and visible test. There was significant decrease in time spent in the target quadrant in animal received testosterone + bicuculline compared to the sham operated group (** P < 0.01) (A). No significant difference was observed in the distance to platform (B). In visible platform test, there were no significant differences in escape latency (C) and velocity (D). T-test (un-paired) analysis was used and P < 0.05 was considered to be statistically significant. (n = 6-8 for each group)
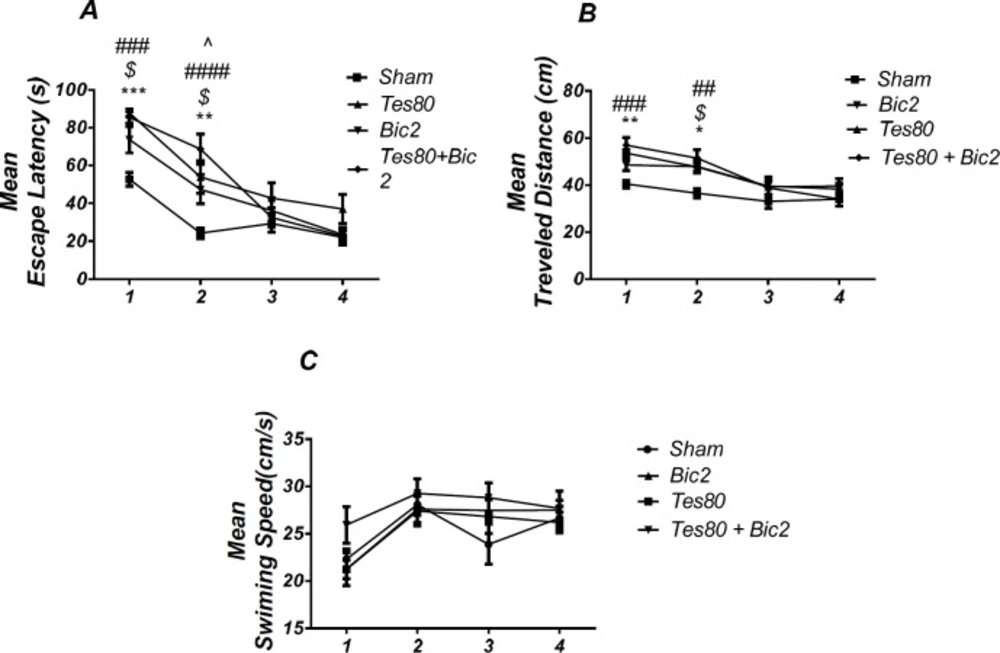
Comparison of testosterone 80, bicuculline 2, testosterone 80 + bicuculline 2 and sham operated group. There was a significant increase in escape latency in1st (*** P < 0.001) and 2 nd (** P < 0.01) days in testosterone treated animals, in 1st and 2 nd ($ P < 0.05) days in bicuculline treated animals, in 1st and 2 nd (### P < 0.001) days in testosterone + bicuculline treated animals compared to the sham operated group. There was a significant increase in escape latency in 2 nd (^ P < 0.05) day in the testosterone + bicuculline treated animals compared to the bicuculline treated animals group (A). Also, there was a significant increase in traveled distance in 1 st (** P < 0.01) and 2 nd (* P < 0.05) days in testosterone treated animals, in 2 nd ($ P < 0.05) day in bicuculline treated animals and in 1 st (### P < 0.001) and 2 nd (## P <0.01) days in testosterone + bicuculline treated animals compared to sham operated group (B). There was not a significant difference in swimming speed in days. There was not a significant difference in swimming speed in days (C). (RM) Two-way analysis of variance (ANOVA) followed by post hoc analysis (Tukey test) were used and P < 0.05 was considered to be statistically significant. (n = 6-8 for each group)
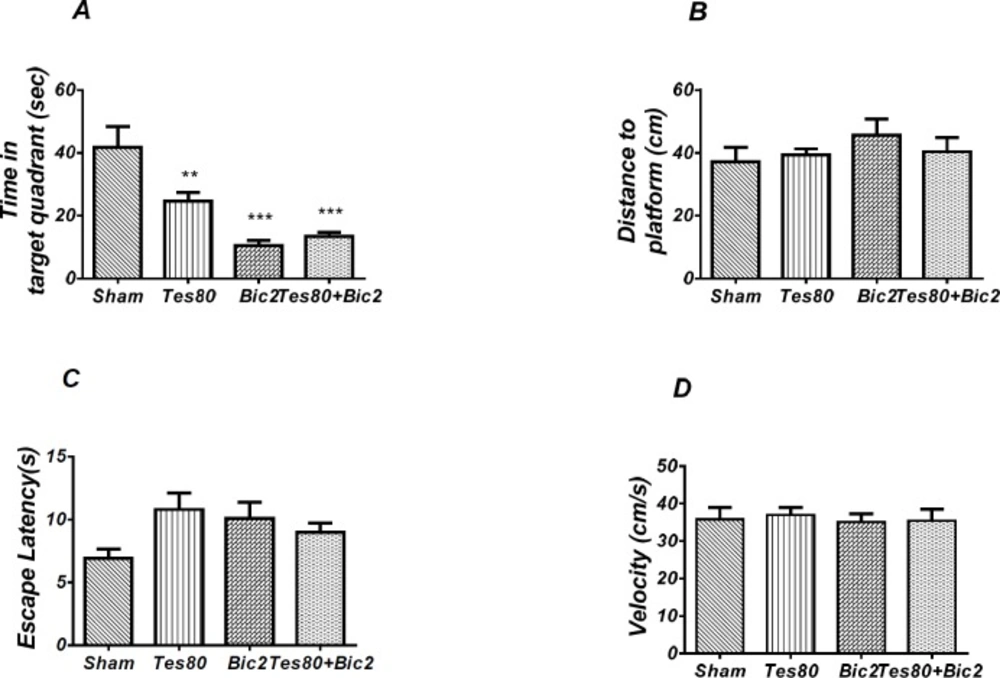
There was decrease significant in time spent in the target quadrant in animal received testosterone, bicuculline and testosterone + bicuculline compared to the sham operated group (** P < 0.01), (*** P < 0.001) (A) and there was no significant difference in distance to platform (B). In visible platform test there were no significant differences in escape latency (C) and velocity (D). One-way analysis of variance (ANOVA) followed by post hoc analysis (Tukey test) were used and P < 0.05 was considered to be statistically significant (n = 6-8 for each group)
