Chemicals, bacterial strains, vectors and DNA techniques
Bacterial growth was done in Luria and Bertani (LB) medium (Merck, Germany) at 37 °C with shaking at 200 rpm. Restriction enzymes were provided from Takara (Shiga, Japan). Human plasminogen and S-2251 were purchased from Sigma Chemicals Company (USA). All chemicals used in laboratory were analytical grade. E. coli DH5a (Stratagene, USA) (f-gyr A96 Nalr, recA1 relA1 Thi-1 hsdR17 r-k m+k) was used as the primary host for the transformation, BL21 (DE3) pLysS (f -ompt hsdB, rB¯ mB¯, dcm gal, DE3, pLYsS cmr) was used to express the SK recombinant protein. The pET-21b (Novagen) was utilized for over-expression of recombinant protein.
Codon optimization, cloning and construction of the Streptokinase expression vector
The codon preferences of E. coli and Streptococcus pyogenes (S. pyogenes) are significantly different. The DNA coding sequence (1323 bp) of Streptokinase from S. pyogenes (GenBank accession no. M19347.1) was taken. Codon optimization was carried out to the codon preference of E. coli genes using NCBI-related database at (http://www.kazusa.or.jp/codon). The optimized SK gene with PelB as a signal sequence, flanked by NdeI and BamHI restriction sites, was synthesized by Shinegene company (China) (GenBank accession no. KT156726.1) then cloned into pUC57 plasmid called pUC-SK. The synthetic PelB-SK was inserted into the pET-21b between the NdeI and BamHI restriction sites. The ligated products were transformed into E. coli BL21 (DE3) plysS competent cells by the CaCl2 method. Screening was done on LB + 100 mg/mL ampicillin and correctness of cloning was confirmed by colony-PCR and sequencing.
Induction and Expression of Recombinant Streptokinase in E. coli DE3
A single colony of E. coli BL21 (DE3) harboring recombinant plasmid (pET21b-PelB-SK) was inoculated in 5 ml of LB medium culture at 37 °C with shaking at 200 rpm overnight and supplemented with 100 mg/mL of ampicillin. To induce the expression of SK protein, 0.5 mM (IPTG) was used when the cell density was reached to OD600 = 0.8 in the shake flask experiments. The expression was done for 12 h.
Isolation of peripelasmic SK, renatuartion of inclusion bodies and molecular weight analysis
The cells in culture were harvested by centrifugation at 4500×g for 10 min at 4 °C and periplasmic expression were obtained by exposing the cell pellet with an equal volume of STE buffer (1 mg lysozyme/mL, 20% (w/v) sucrose, 30 mM Tris/HCl (pH 8.1), 1 mM EDTA) on ice for 10 min. Cell debris was separated by centrifugation according to our previous paper (
12).
The separated periplasmic SK was as insoluble inclusion bodies, so it needed a renaturation to reach an active form of recombinant enzyme. First the inclusion pellets were solubilized in 8 M urea buffer at pH 8. The mixture was incubated at 25 °C for 1 h before the insoluble parts were removed by centrifugation. The solution was then diluted with phosphate buffer (pH 10.7) for SK renaturation. The solution was dialysis against the buffer [20 mM Tris/HCl pH 8.0, 50 mM NaCl, 1 mM EDTA] at 4 °C overnight.
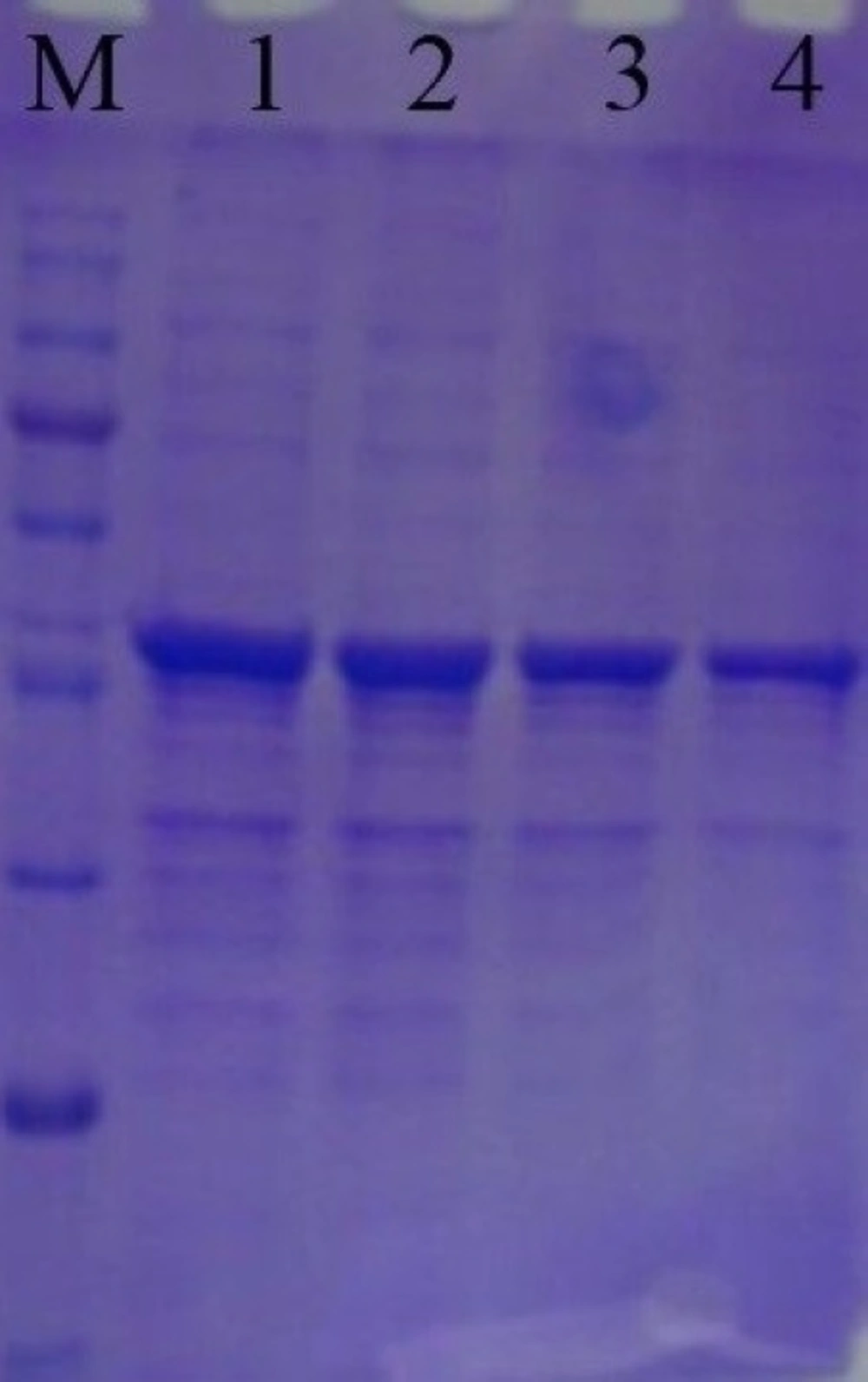
Recombinant SK protein was analyzed by 15% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). The protein band of recombinant SK was visualized by Coomassie Blue staining. The amount of total protein was determined by the Bradford method, using bovine serum albumin (BSA) as a standard.
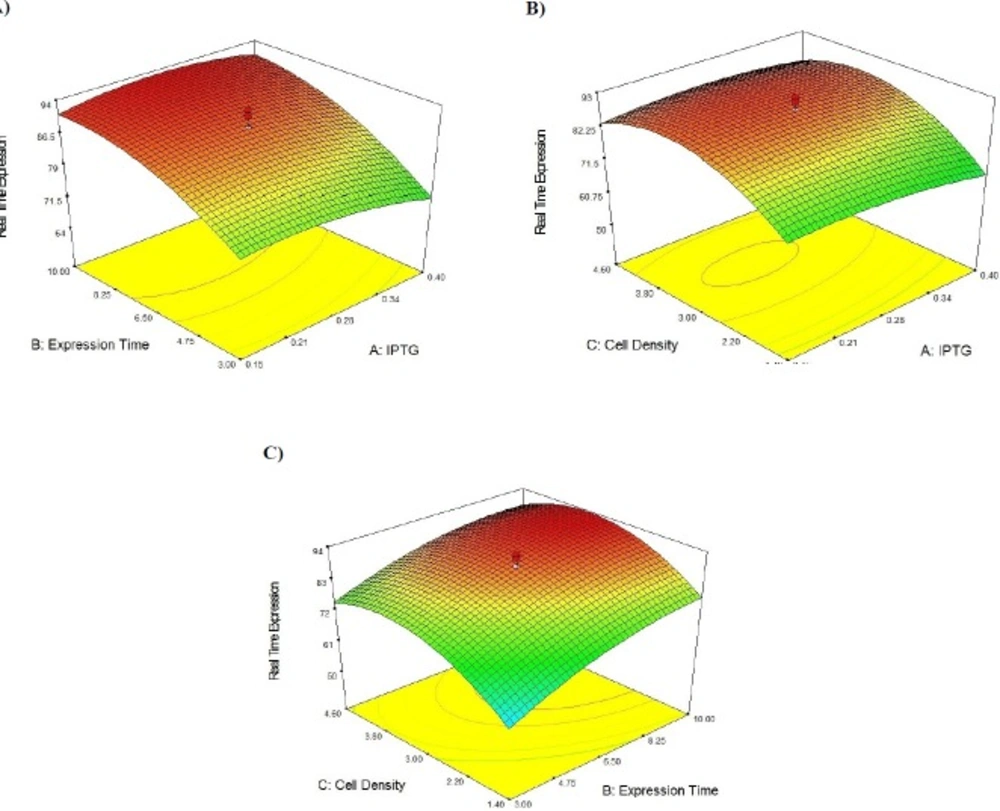
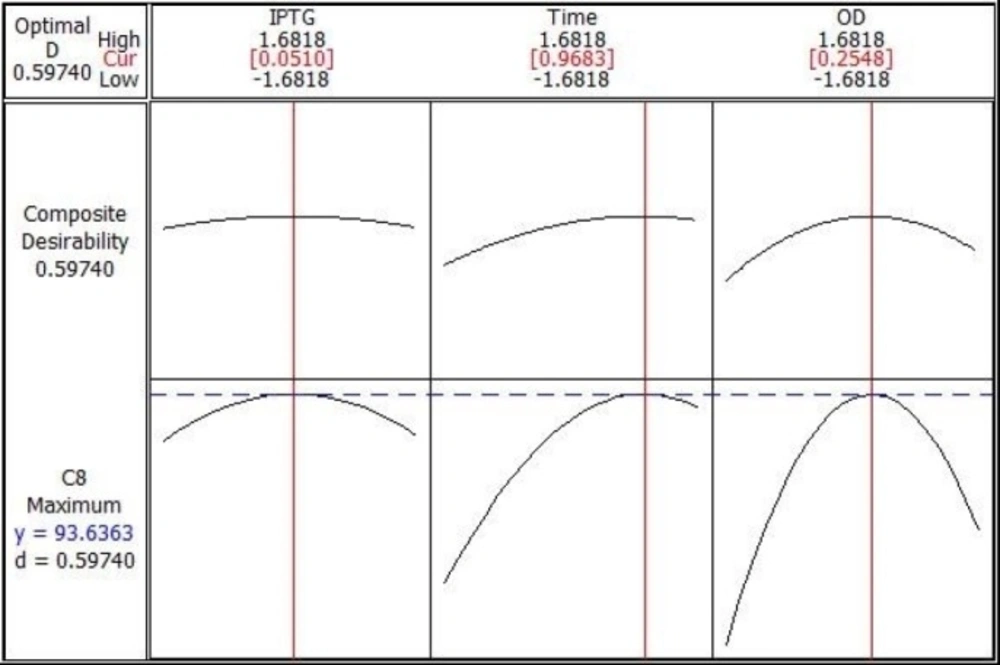
Overexpression of recombinant Streptokinase using response surface methodology
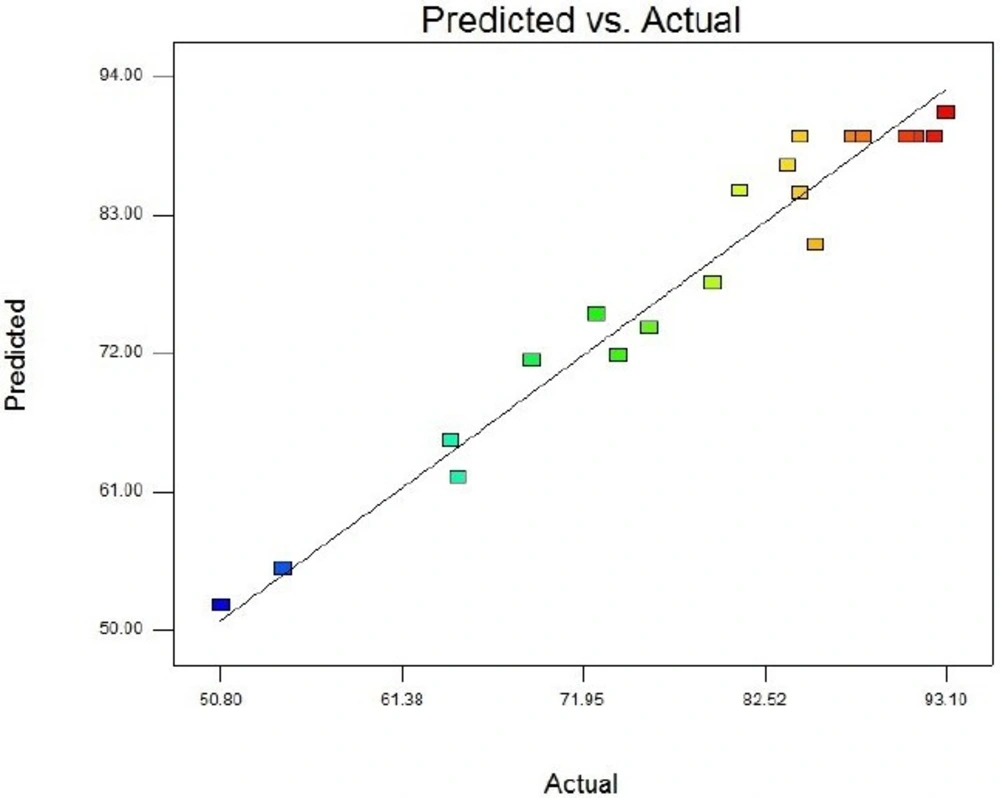
Central Composite Design (CCD) and Response Surface Methodology (RSM) were used for optimization of recombinant Streptokinase expression level in E. coli. Minitab 16 software (Minitab Inc., USA) was taken to design the experiment and optimization of three significant cultivation conditions: IPTG concentration, post-induction period and cell density of induction (OD600). Twenty experiments contain six replicated center points were designed and carried out. The analyses of experimental data were carried out statistically by regression method:
Y = β0 + ∑βi xi + ∑βiixii 2 + ∑βijxi xj + ε
Where Y is the predicted streptokinase mRNA percentage, β0 is a constant coefficient, βi the linear coefficient, βii the quadratic coefficient and βij the cross-product coefficient. Xi and Xj are input independent variable levels, while ε is the residual error. Design Expert software 7.0.0 was employed for Data analysis of experimental design and surface response methodology. The transcriptional level of recombinant Streptokinase was measured by Real Time PCR method in different conditions. Recombinant E. coli host cells transformed with pET21b-PelB-SK vector without induction were used as negative control.
Quantitative analyses by real-time PCR and ΔΔCt method
Total RNA from E. coli BL 21 cells contain recombinant SK were isolated using Trizol reagent (Life Technologies, USA) following the standard protocol. First, the cells were homogenized in 1 mL Trizol solution and 200 μL chloroform was added to samples and mixed completely for 3 min. Then, the mix was centrifuged at 12,000×g for 15 min at 4 °C. The upper aqueous phase was transferred carefully into new tube without disturbing the interphase and equal volume of isopropanol was added in the tube. The mixtures were thoroughly resuspended and incubated in -20 °C for 30 min and centrifuged at 12,000 ×g for 10 min at 4 °C. The precipitated RNA pellets were washed with 1 mL ethanol (75%, v/v). RNA pellets were recovered after centrifugation at 12,000 ×g for 5 min at 4 °C. RNA samples were allowed to air-dry for 2–3 min and then resuspended in 30 mL diethyl pyrocarbonate-treated water (Life Technologies, USA). The extracted RNA was quantified by measuring absorbance at 260/280 nm by Nanodrop and the quality of RNA purified was checked by gel electrophoresis. cDNA synthesis was carried out with Thermo Scientific cDNA synthesis kit. Reverse transcription was followed using 50 mg total RNA (maximally in 20 mL) and 1 mL random hexamer primers. The volume of the assay mixture was adjusted to 12 mL with RNase-free water, and then the mixture was incubated for 5 min at 70 °C, followed by incubation for 10 min at room temperature to allow the primers to anneal with the RNA.
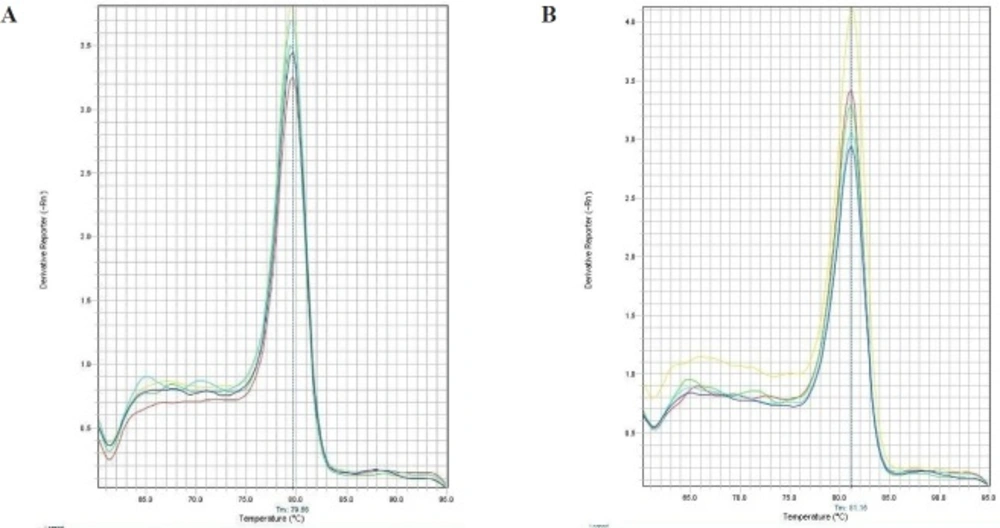
For analysis of streptokinase expression level, Real Time PCR was performed by using Power SYBR® Green PCR Master Mix (life technology), according to manufacturer’s instructions (Applied Biosystems, USA). The kit has a Hot-start
Taq DNA polymerase. All samples were analyzed in duplicate and the average value is reported. For determination of the mRNA level, 16S rRNA was used as internal control gene. The primers used for this study were designed using Web-based Oligo7 Primer Analysis Software. The primers for streptokinase were: sense 5- CATAAACTGGAAAAAGCCGATCTG -3 and antisense 5- GACCGCTCAGCAGAAATTCTTG -3. The primers for 16S were: sense 5- CTACGGGAGGCAGCAGTGG -3 and antisense 5-TATTACCGCGGCTGCTGGC -3. The StepOne™ Real-Time PCR Systems (ABI) was used to detection relative quantification. The amplification reactions were done under following conditions: 10 min at 95 °C, followed by 45 cycles at 95 °C for 15 sec, 60 °C for 1 min. Melting curve program was set to 60-95 °C with a heating rate of 0.1 °C per second and a continuous fluorescence measurement. In order to identify the specificity of amplification products a dissociation curve was plotted (
Figure 1). The 2
-ΔΔCt method was used to analyze the relative changes in the level of gene expression (
13).
Determination of Streptokinase activity
SK activity was assayed using by chromogenic substrate method that was an endpoint method. The Streptokinase transformed plasminogen to plasmin in solution, in the existence of chromogenic substrate S-2251 (H-D-valyl-L-leucyl-L-lysine-p-nitroanilide dihydrochloride; (Sigma, USA), in the absence of fibrin.
Substrate solution included a mixture of 1 mL of 0.5 M Tris–HCl pH 7.4, 1 mL of 3 mM S- 2251 and 5 μL of 10% Tween 20. This solution was kept at 37 °C and immediately before use, 45 μL of human-plasminogen solution (1 mg/mL) was also added to substrate solution. Streptokinase solution tested at different concentrations for the dose-response curve. Streptokinase diluted in 10 mM of Tris–HCl (pH 7.4) at 37 °C, 0.1 mM NaCl and 1 mg/mL albumin to reach 4.0, 2.0, 1.0, 0.5 IU/mL concentrations and maintained at 37 °C in a microtiter plate. In the test samples, the reaction was performed after addition of 60 μL of Streptokinase solution to 40 μL of substrate solution. The absorbance of the wells was measured at 405 nm for 20 min before an endpoint OD was taken immediately.