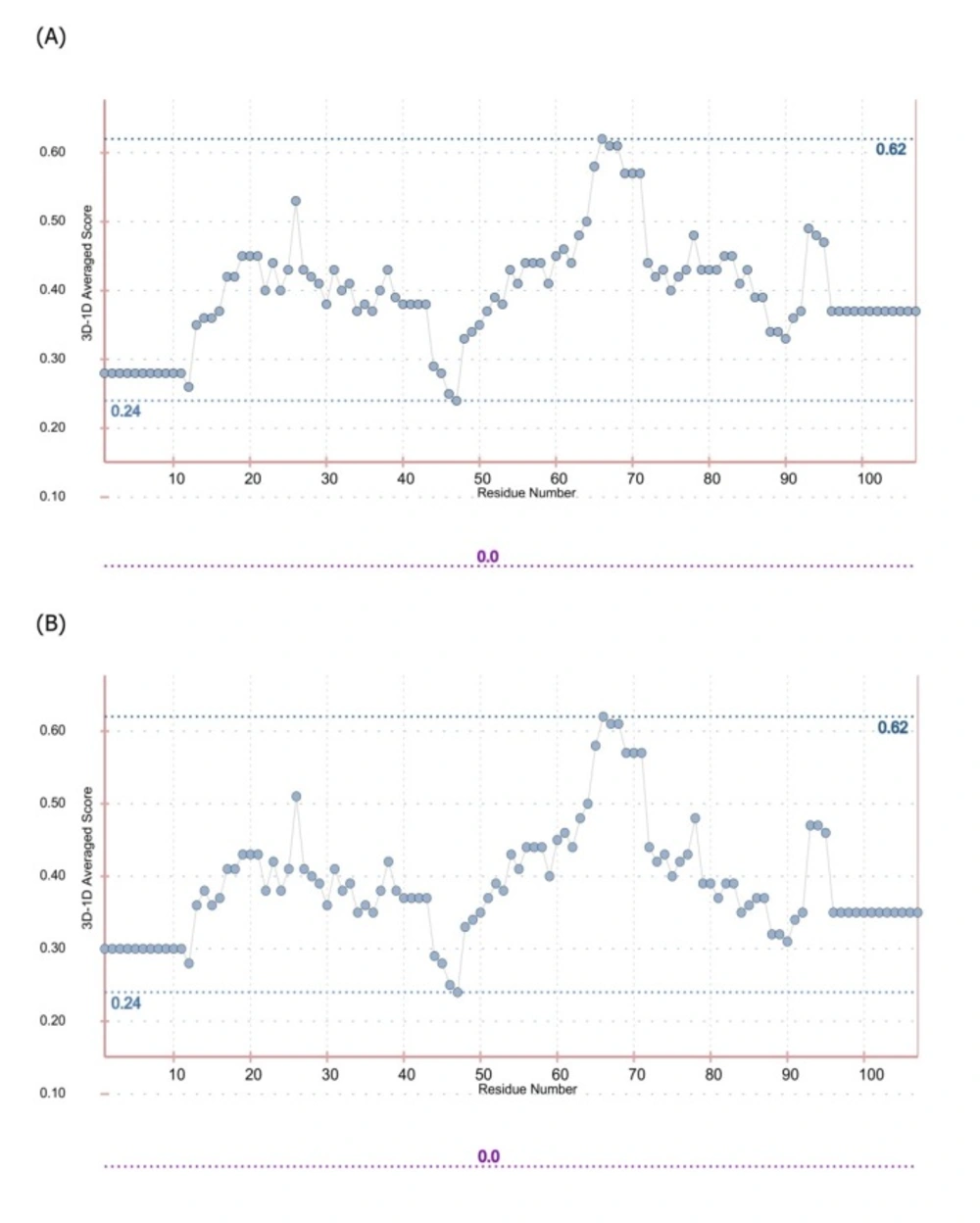
Progress of inflammatory conditions greatly depends on TNF-α aberrant production. Therefore, prevention of TNF-α activity would result in promising outcomes in patients suffering from inflammatory diseases. In this regard much attention has been devoted to develop anti-TNF-α agents which could directly bind to TNF-α suppressing its adverse effects. These inhibitory molecules vary from small molecules such as peptides to macromolecules such as full length antibodies (
34-
42). Up to now, only few full length anti-TNF-α antibodies have been approved by FDA to be used in inflammatory diseases (
12). Antibodies are biological agents that are extensively used in clinic and diagnosis due to their high specificity and affinity towards corresponding antigens (
20). Despite having the advantage of specificity, the immunogenicity and pharmacokinetic problems as well as the time consuming and expensive process of production of such therapeutic antibodies are challenging issues (
43-
48). Therefore, identification of small format of antibodies which provide manageable physicochemical properties with suitable specificity towards target of interest is of great importance. Such attitude paved the way for introducing the therapeutic Fab antibodies such as abciximab (an anti-PIIb/IIIa), ranibizumab (an anti-VEGF), and certolizumab pegol (an anti-TNF-α) (
49-
51). Tomlinson I&J phage display antibody libraries are non-immunized human phage libraries composed of scFv displaying phagemids developed by Greg Winter and colleagues in 2000 (
52). Although Tomlinson phage libraries are designed based on scFv format, in our panning process against TNF-α, two antibodies namely, I44 and I49, possessing only one domain of scFv antibodies (
i.e. V
L) were identified (
23). Compared to larger formats of antibodies such as full length, Fab, and scFv antibodies, domain antibodies (dAbs) are more stable non-aggregating molecules, which make them suitable tools for many purposes such as inhibition of cytosolic/nuclear proteins that cannot be targeted with genetic knockout techniques due to the much easier folding of dAbs inside the cytosol (
53). On the other hand, because of the small size, domain antibodies can be administered not only by injection but also from oral route. These excellent features of dAbs compared to complete antibodies and scFv fragments have attracted much interest of different companies around the world such as Ablynx, Domantis (GSK), Ossianix to investigate on identification of new dAbs for therapeutic purposes. Ozoralizumab, produced by Ablynx company for rheumatoid arthritis, is anti-TNF-α single domain antibody which has successfully passed phase IIa clinical trial study. Furthermore, several therapeutic domain antibodies are in different phases of preclinical and clinical trials in Ablynx, Domantis (GSK), and Ossianix companies. For example, caplacizumab (ALX-0081) is the first single domain anti-von Willebrand factor antibody which is going to be marketed for treatment of thrombotic thrombocytopenic purpura and thrombosis by Ablynx Company (
53). Based on this evidence, we aimed to produce and purify the previously identified single domain anti-TNF-α antibodies namely, I44 and I49, in order to investigate their binding to and inhibition of TNF-α. To do so, phage particles harboring the coding gene for I44 and I49 were used to infect
E.coli origami (DE3) bacteria. Presence of PelB signal peptide in phage genome enables periplasmic expression of the target proteins (
i.e., I44 and I49). This feature provides an advantage for the current expression system where periplasmic compartment supplies a reduced environment suitable for the formation of disulfide bonds in 3D structures of I44 and I49 proteins (
54,
55). This expression and purification system resulted in successful production of I44 and I49 antibodies with high purity, evidenced by SDS-PAGE and western blotting analyses (
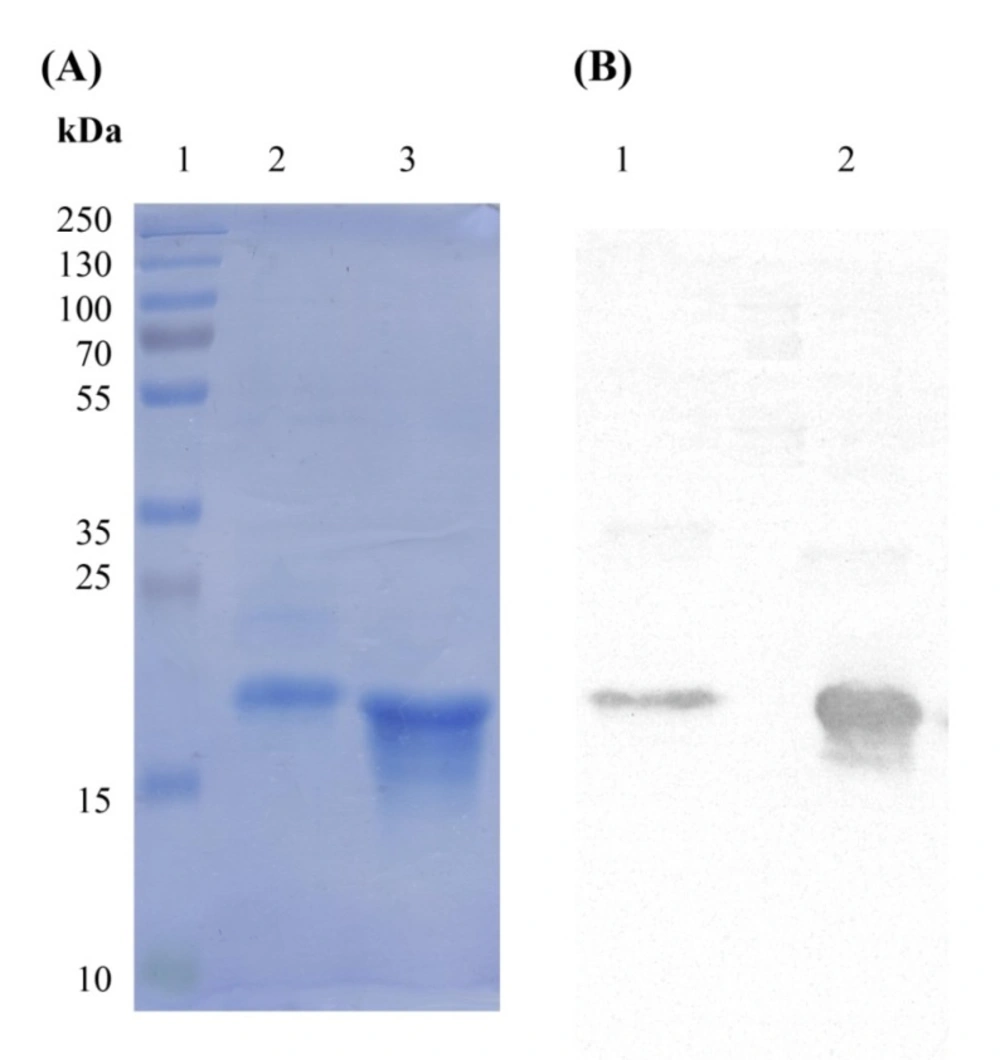
Figure 2). The variation in the amount of produced I44 and I49 proteins is understandable as few differences in amino acid sequences of proteins can result in much variability in protein production in the same condition (
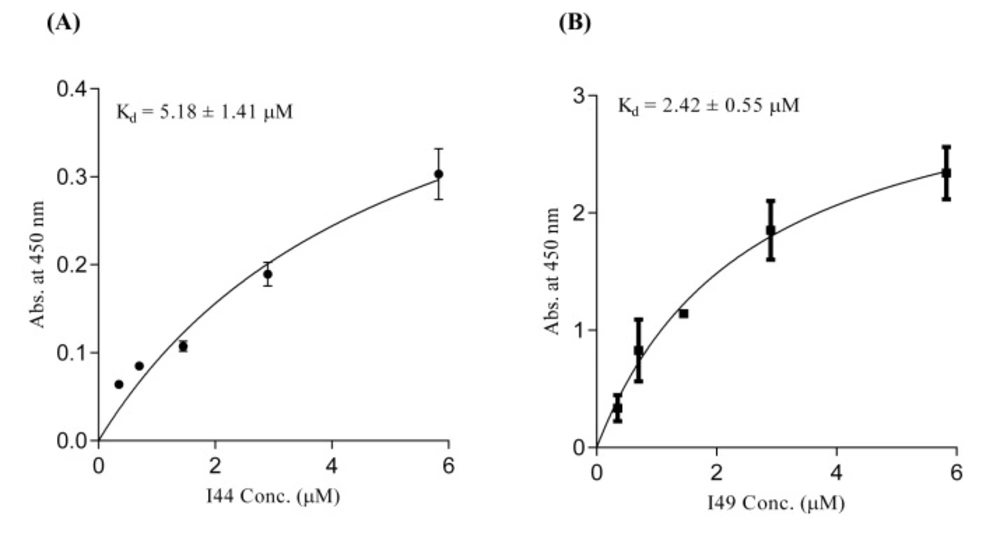
56). In ELISA experiment I44 and I49 showed appropriate affinity towards TNF-α with K
d values of 5.18 ± 1.41 and 2.42 ± 0.55 µM, respectively, which are comparable with results reported elsewhere for small format anti-TNF-α antibodies such as scFvs (
57,
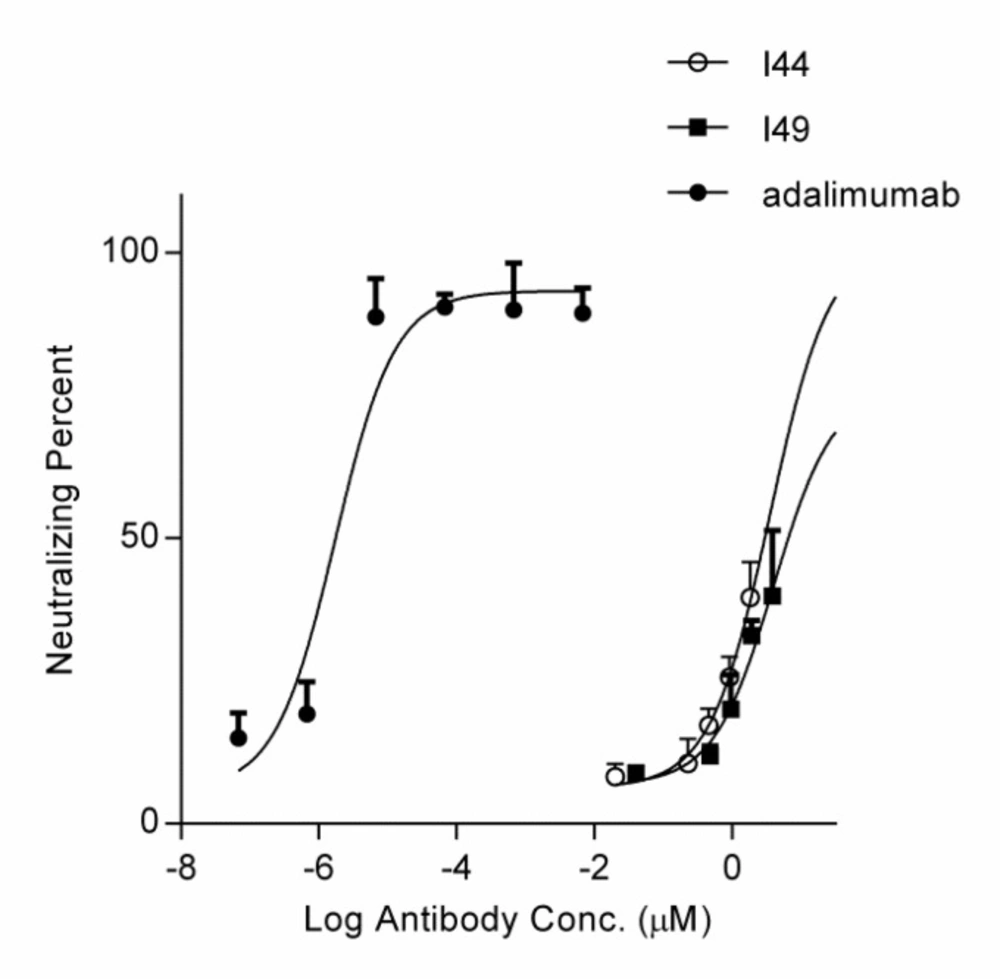
58). The inhibitory effect of I44 and I49 antibodies on TNF-α biological activity was evaluated by MTT assay where TNF-α sensitive L929 cells were survived from TNF-α cytotoxic effects in the presence of different concentrations of the dAbs. The determined IC
50 for I44 and I49 were 6.61 and 3.64 µM, respectively which are much bigger than that of adalimumab (1. 67 ± 0.32 pM). Such big difference in IC
50 values can be attributed to the lower binding valence of I44 and I49 single dAbs (~17 kDa) compared to adalimumab (148 kDa). To identify the most important residues involved in antibodies-TNF-α interactions, molecular docking was carried out in which the modeled structures of dAbs were docked onto TNF-α trimer. The docking scores obtained from ZDock program were 952.23 and 981.76 for I44-TNF-α and I49-TNF-α complexes, respectively, which is in close agreement with the data achieved from ELISA experiment where I49 showed higher affinity towards TNF-α compared to I44. In the next step, the possible interactions between dAbs and TNF-α were predicted using PIC web server (
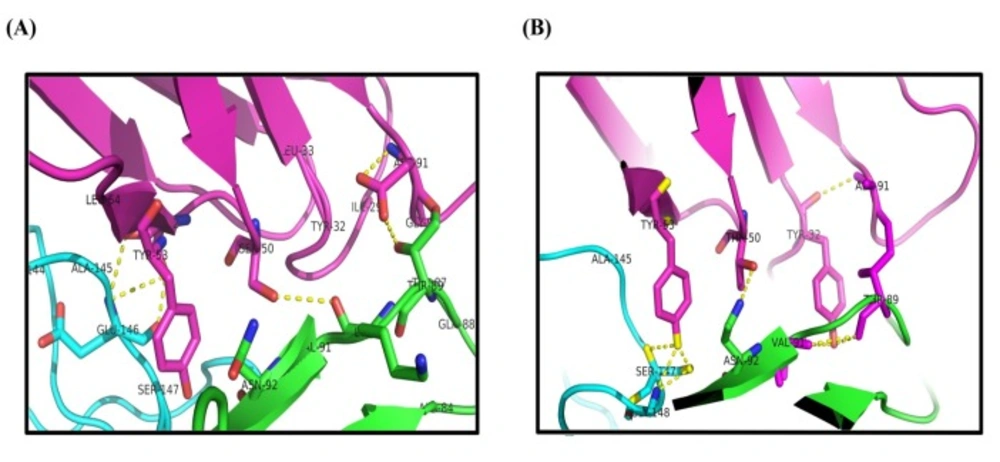
Table 3). According to PIC analyses, the main hydrophobic contacts within 5 Å between dAbs and TNF-α are established via Tyr
32 in CDR1 of both antibodies and Tyr
87 and Val
91 of TNF-α (
Figure 7). In addition, Tyr
53 and Leu
54 located at CDR2 of dAbs are also contributing in hydrophobic interactions involving Leu
143 and Ala
145 of TNF-α (
Figure 7). These results are in close agreement with the crystallographic data of adalimumab-TNF-α complex where the hydrophobic interaction of adalimumab with Val
91 of TNF-α has been elucidated (
59). Apart from hydrophobic contacts, dAbs-TNF-α complexes have been stabilized by hydrogen bonds. The most important hydrogen bonds are detected between I44 Ser
50 and I49 Thr
50 at CDR2 of antibodies and Asn
92 of TNF-α as well as I44 Tyr
32 (CDR1) and I49 Tyr
53 (CDR2) with Val
91 and Gly
148 of TNF-α, respectively (
Figure 7). Such intermolecular hydrogen bonds are observed in the crystal structure of adalimumab-TNF-α complex (
59). Comparison of the possible interactions of these dAbs with the interactions of adalimumab and infliximab with TNF-α revealed that the interaction mode of I44 and I49 is similar to adalimumab rather than infliximab. It has been shown that four distinguished clusters of TNF-α consisting of residues ranging from 20-23, 65-92, 110-115, and 135-146, are participating in adalimumab binding (
59). Comparing this information with the data obtained from molecular docking study indicates that the identified dAbs mimic the binding mode of adalimumab, interacting with TNF-α residues ranging from 84-92 and 143-148. However, no interactions between the dAbs and TNF-α residues ranging from 20-23 and 110-115 were observed which can be reasoned for their low effectiveness compared to adalimumab. Therefore, structural modifications on the identified dAbs for designing more potent anti-TNF-α antibodies are required. This goal can be achieved by altering the key residues of the antibodies responsible for TNF-α binding in order to determine the role of each residue in TNF-α binding to propose mutations for improvement of TNF-α binding characteristic of the designed antibodies. Moreover, TNF-α binding abilities of the dAbs, identified in the current study, can be refined by pairing these antibodies with a library of heavy chain phage libraries (chain-shuffling) (
60,
61).