Cultivation of L. major parasites
The promastigotes (MRHO/IR/75/ER) were cultured in RPMI-1640 medium (Gibco-BRL, USA) and L-glutamine (20 mM), containing 10% fetal bovine serum (FBS; Gibco-BRL, USA), 100 μg/mL of streptomycin (Sigma), and 100 U/mL of penicillin; incubation was then carried out at 26 °C. Using an inverted microscope, the growth of
L. major promastigotes was examined each day (
36).
Culture of Raw 264.7 macrophage cells
To evaluate cytotoxicity and obtain an amastigote macrophage culture, the RPMI-1640 medium, containing streptomycin (100 μg/mL), 10% FBS, and penicillin (100 U/mL), was used for culturing at 37 °C (5% CO2). The macrophage cell growth was evaluated daily using an inverted microscope. When the cells reached about 80-90% confluence, they were subcultured into a new flask containing fresh cell culture medium.
Drug preparation
Oxaliplatin was purchased from Sigma Aldrich Co. (USA) as white powder with 99% purity and a molecular weight of 397.29 g/mol. After dissolving oxaliplatin in distilled water (1 mL) to obtain a 5-mg/mL stock concentration from the drug, it was aliquoted in 0.5 mL vials (Eppendorf tubes) for storage at 4 °C until use. For in-vitro experiments, different concentrations of drugs (i.e., 400, 200, 100, 50, 25, 12.5, 6.25, 3.12, 1.56, and 0.78 µg/mL) were prepared using the RPMI-1640 medium.
Anti-promastigote activity assay, proliferation assay
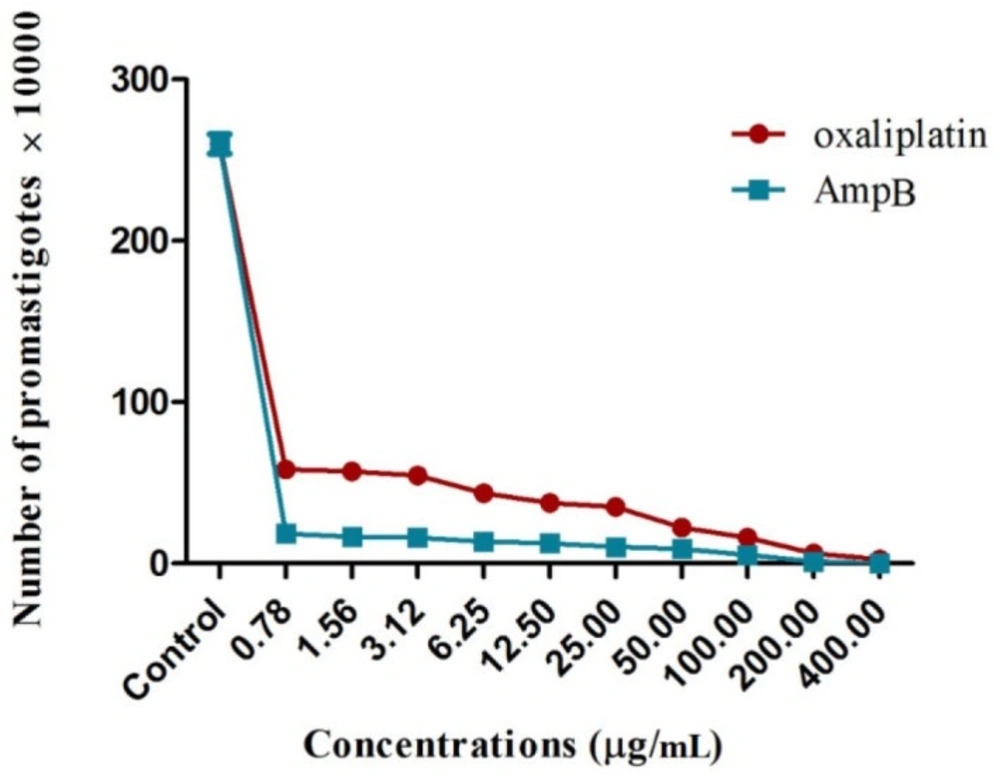
To examine the anti-leishmanial effects, L. major promastigotes in the logarithmic growth phase (2 × 106 parasites/mL) were cultured in 96-well microtiter plates containing RPMI-1640 medium with 10% FBS; they were then incubated at 24 ± 2 °C overnight. Following incubation, 100 µL of different oxaliplatin and AmpB concentrations were used to treat the promastigotes for 72 h. The promastigotes cultured with no drugs and RPMI-1640 medium with no promastigotes or drugs were respectively used as the negative control and blank; the experiments were repeated three times. The number of promastigotes was counted under an optical microscope to determine the anti-leishmanial effects, using a hemocytometer chamber. In addition, IC50 was assayed by non-linear regression analysis.
Promastigote viability test
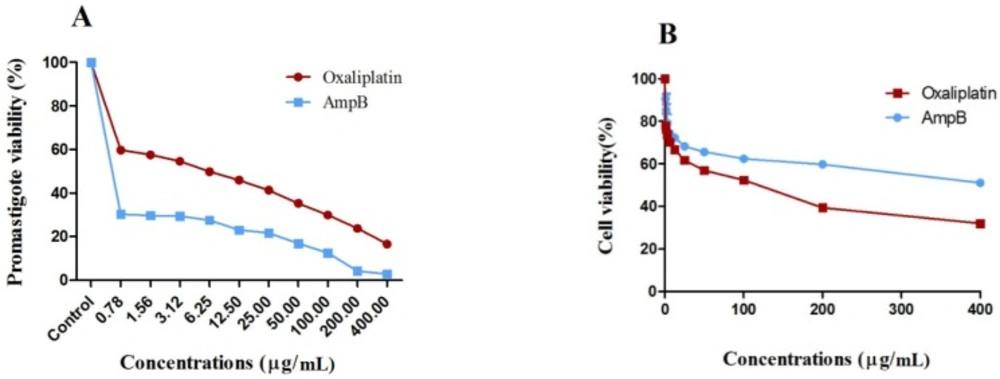
The L. major promastigotes were evaluated in terms of mitochondrial oxidative activity by measuring parasite viability with the colorimetric MTT method (Sigma Aldrich, USA). Briefly, L. major promastigotes (2 × 106 cells/mL) were cultured in plates (Nunclon®, Denmark) containing oxaliplatin (400 to 0.78 µg/mL) at 24 °C for 72 h. On the other hand, the positive control was AmpB, and the negative control included promastigotes cultured in RPMI-1640 medium with 10% FBS without the drug.
Incubation was performed for three hours at 37 °C after adding 5 mg/mL of MTT solution (20 µL) to the wells. Then, DMSO (100 µL/well) was used to solubilize formazan crystals at room temperature for half an hour (
37). An ELISA reader (ELX800) was used to read absorbance at 570 nm. In addition, the IC
50 of
Leishmania growth was measured by applying non-linear regression analysis (
38). The following formula was measured to determine the viability percentage of promastigotes (
39).
[(AT–AB)∕(AC–AB)] × 100%
where AB, AC, and AT represent the OD of the blank well, negative control, and treated cells, respectively.
Cytotoxicity and selectivity index
Macrophage viability was determined using the MTT method (Sigma Aldrich, USA). Briefly, after seeding murine macrophage Raw 264.7 cells in 96-well microtiter plates in triplicate, incubation was performed overnight at 37 °C. Then, a fresh medium (100 µL) was used to replace the medium. The cells were treated using different oxaliplatin concentrations as serial dilutions, and AmpB was used as the positive control at 37 °C. After incubation for 72 h, 5 mg/mL of MTT solution (20 µL) was poured in the wells, and incubation was repeated at 37 °C for three h.
The formazan crystals were solubilized in DMSO (100 µL/well) for 30 min at room temperature. The colorimetric reaction was read using an ELISA reader (ELX800) at 570 nm. The non-linear regression analysis was used to measure the CC
50 value in GraphPad Prism 6.07 (GraphPad Inc., USA). The selective index (SI) was also calculated by measuring the CC
50 to IC
50 ratio (
38).
Anti-amastigote activity
After seeding murine macrophage-derived Raw 264.7 cells on round glass coverslips in 12-well plates containing RPMI-1640 medium (with 10% FBS), they were incubated at 37 °C overnight. The adherent macrophages were exposed to L. major promastigotes (macrophage-parasite ratio1:10) after washing the non-adherent cells with phosphate buffered saline (PBS); they were then placed at 37 °C for 24 h for phagocytosis of promastigotes.
For the removal of free promastigotes, the infected macrophages were washed in PBS. For treatment, 6.25, 12.5, and 25 µg/mL of oxaliplatin and AmpB as the positive control were used at 37 °C. The medium with the infected macrophages but without the drug was used as the negative control. After 72 h, the coverslips were fixed in methanol and stained with Giemsa stain solution. The percentage of infectivity was assessed for each sample by measuring the ratio of infected macrophages to 300 macrophages in triplicate. The infection index rate (IIR) represents the mean amastigote count per macrophage cell (
40,
41).
Detection of phosphatidyl serin exposure
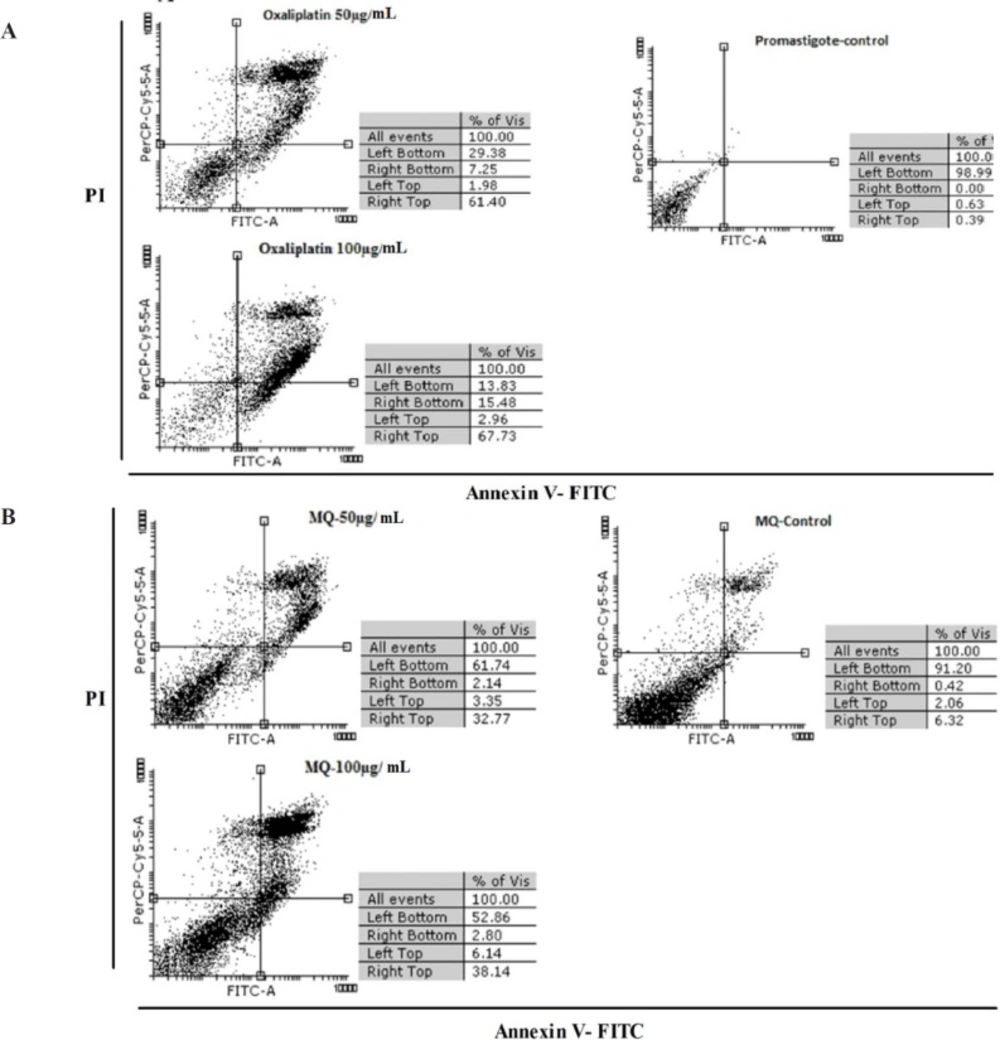
An Annexin-V FLUOS staining kit (Bio-vision, USA) was used to evaluate oxaliplatin induced apoptosis. In brief, L. major promastigotes in murine macrophage derived Raw 264.7 cells were exposed to 50 and 100 µg/mL concentrations of oxaliplatin, as mentioned above. Promastigotes and macrophages exposed to no drugs were used as the control groups. After 72 h, the cells were washed in cold PBS and harvested by centrifuging at 14,000 g for 10 min.
The supernatants were removed and the pellets were resuspended in 500 μL binding buffer, 5 μL annexin-V, and 5 μL propidium iodide (PI). Then, the samples were incubated in darkness at 26 °C for 15 min. Apoptosis was assayed by detecting the PI fluorescence intensity using a FACS Calibur flow cytometer (Becton Diekinson). The results were analyzed using CellQuest software.
Experimental animals
In the present study, Razi Institute of Iran provided female BALB/c mice (4-6 weeks old; 18-20 g). They were kept under optimal conditions (25-26 °C; relative humidity, 55-65%) at the animal facilities of Tarbiat Modares University, Tehran, Iran. The ethics committee of the university approved all animal procedures. We used 36 mice which were infected subcutaneously in the tail base using stationary phase promastigotes. When the nodules were developed in the injection site, the animals were randomly divided into three groups (12 mice per group). Then, animals of each group were divided into three subgroups (n = 4 mice) to measure lesion size, parasite burden, and cytokine assay.
Development of leishmanial lesions and treatment
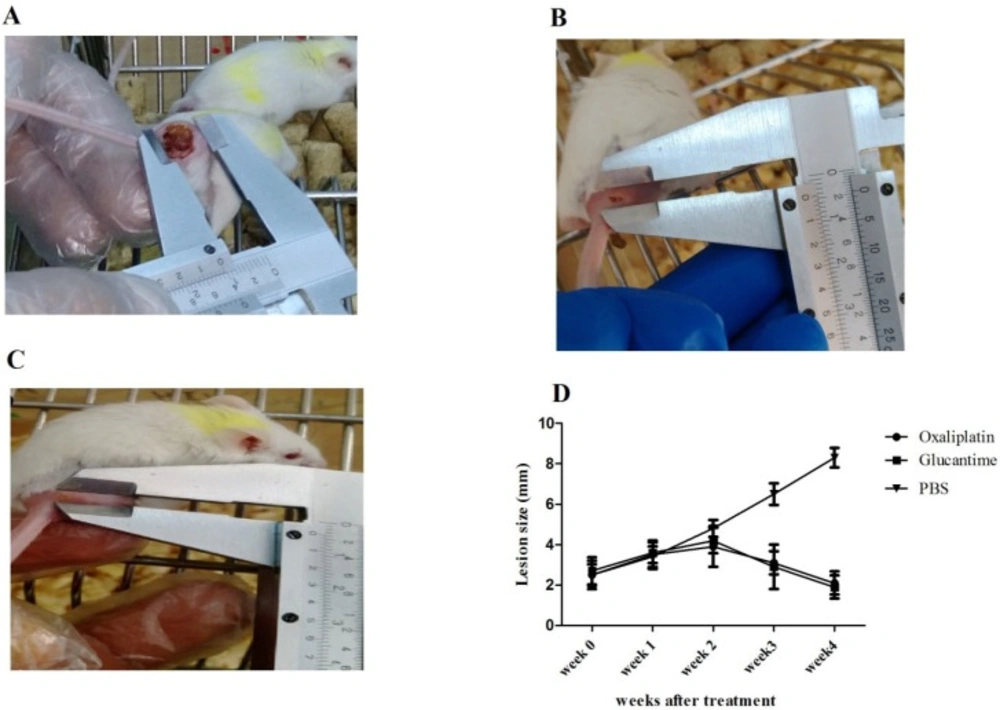
In this study, the mice were infected subcutaneously in the base of the tail using 100 µL of a stationary phase promastigote suspension (1 × 106 parasites). The animals were divided into groups when the nodules developed in the injection site (15-20 days after infection). Group I received 25 mg/kg of oxaliplatin daily for 30 days (i.p. route), group II received 20 mg/kg of glucantime daily for 30 days (IM route), and group III (control) received 200 µL of PBS (i.p. route). From each group, four mice were selected to measure the lesion size. To examine the drug efficacy, the width and length of the lesion were weekly measured using a digital caliper. The differences in the mean lesion size were assayed by Two-way ANOVA and Bonferroni post-hoc tests.
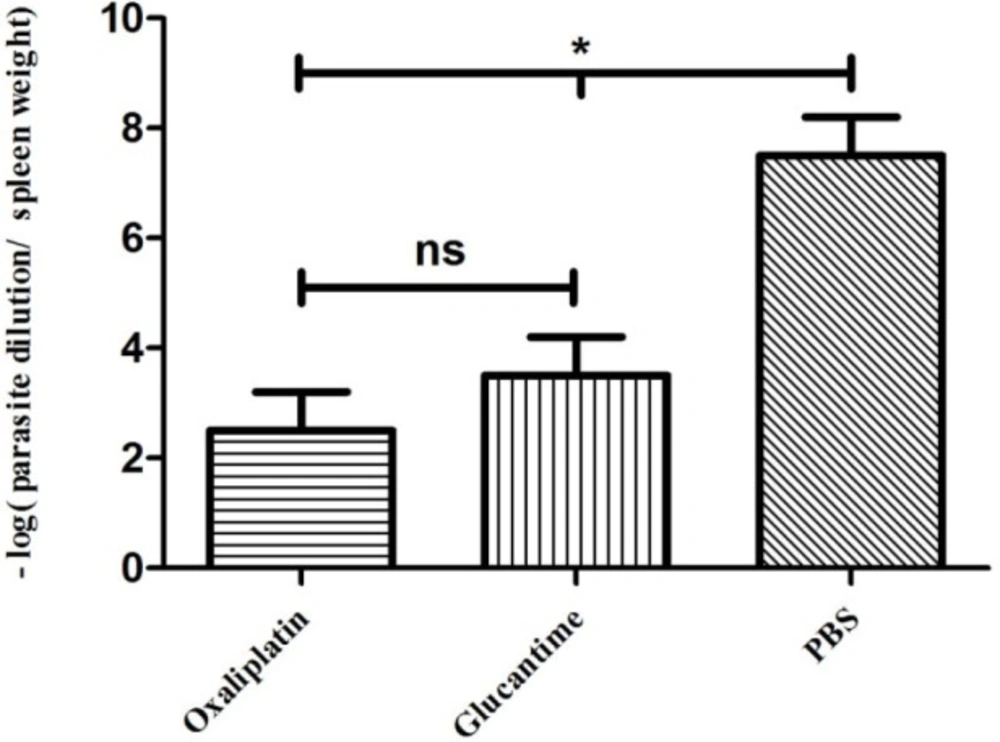
Parasite burden
To determine the parasite burden, four mice were euthanized from each group at four weeks post-treatment, and their spleens were removed under aseptic conditions.
The homogenized spleen tissue (10 mg) was cultured in 96-well cell plates containing RPMI-1640 medium with 100 IU/mL of penicillin, 10% FBS, and 100 µg/mL of streptomycin at serial dilutions of 1-10-20. Then, incubation was carried out at 26 °C. After seven and 14 days, to determine mobile promastigotes in each well, the plates were evaluated using an inverted microscope at 40X magnification. The final dilution with at least one motile promastigote was considered as the final titer. The following equation was used to determine the number of parasites:
parasite burden = -log (parasite dilution/tissue weight).
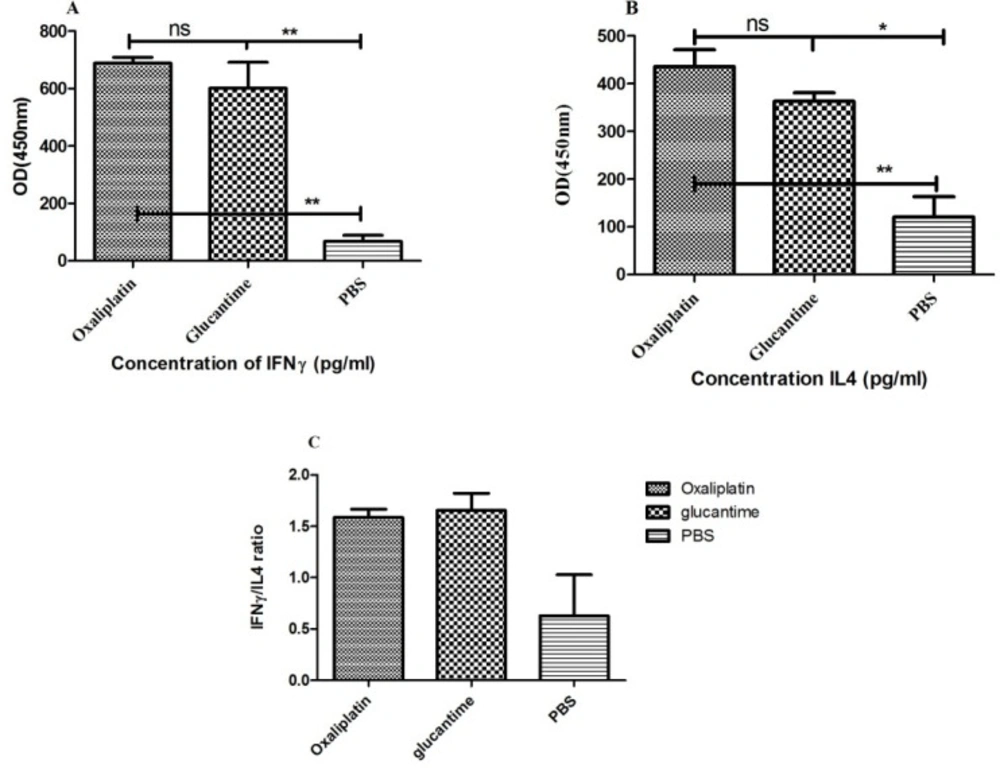
Cytokine assay
For IFN-γ and IL-4 measurements, splenocytes were exposed to soluble
Leishmania antigen (SLA), obtained from stationary phase promastigotes according to previous studies (
42). After euthanizing four mice from each group at four weeks post treatment, the spleens were extracted and aseptically homogenized in PBS.
The supernatant was removed after double washing in PBS, and the cell pellets were resuspended in cold ammonium-chloride-potassium lysis buffer (5 mL; 0.15 M NH
4Cl, 10 mM KHCO
3, and 0.1 mM Na
2EDTA); also, erythrocytes were lysed by incubating for 5 min at room temperature. After washing, 3 × 10
6 cells/well were seeded in the plates, containing RPMI-1640 medium, which was supplemented with 100 μg/mL of streptomycin, 10% FBS, and 100 U/mL of penicillin; they were then exposed to 25 μg/mL of SLA at 37 °C. The supernatant was harvested after 72 h to assay cytokine levels (
43). Also, ELISA kits (MABTECH, USA) were used, as outlined by the manufacturer, to measure IL-4 and IFN-γ.
Statistical analysis
For data analysis, GraphPad Prism 5.0 (Graphpad Inc., USA) was used. The results were compared using parametric tests including unpaired samples t-test and One-way or Two-way ANOVA. p-value < 0.05 was considered significant. The values are presented as mean ± SD. Graphpad Prism was used for plotting the graphs.
Ethical considerations
The Ethics Committee of Tarbiat Modares University approved all stages of the study, including animal maintenance and handling (ID.IR.TMU.REC.1394.259).