Induction of the hairy roots
The fresh leaves of
Atropa komarovii plantlet with 6 or 7 leaves demonstrated high affectability to ATCC15834 (
Figure 1). The high change recurrence was almost seen with 60% from leaf explants vaccinated by means of
A.rhizogenes and other
Atropa species such as
belladonna and baetica have been genetically modified by
Agrobacterium rhizogenes (
14) The hairy roots and control roots were extracted from explants and exchanged to new MS medium with no auxin, and the hairy roots, fit for blending endogenous auxin, in this manner did not require exogenous auxin. After one month, some control roots were not survived but rather changed roots achieved greatest biomass, which had lots of horizontal expanding, thick, negative geotropism, and stretched quickly (
Figure 1, A,B,C). Tissue culture, joined with genetic designing particularly changed innovation that caused fulfillment and opened new ways for high volume generation of pharmaceutical substances (
15). This examination demonstrated an effective technique for prompting hairy roots in
Atropa komarovii. The outcomes introduced in this reveal that the utilization of
A.rhizogenes may be a fruitful way to deal with hairy root infusion (
16). In the contamination method, the initial phase is the host/pathogen interface where a few components commit to the foundation of this connection. Phenolic compounds and sugars secreted from the plants, according to plant genotype, thus created distinctive reactions (
17).
Agrobacterium rhizogenes intervened genetic change in plants is a settled auxin: cytokinin proportion in the plant cell. Since different plants can demonstrate discriminative susceptibility to a given
Agrobacterium rhizogenes strain.
A. rhizogenes that was harboring double vector was developed with leaves of
A.komarovii approximately following three week of incubation on MS basal medium. The roots rose straightforwardly from the site of contamination on leaf discs, at the injured locals (
18).
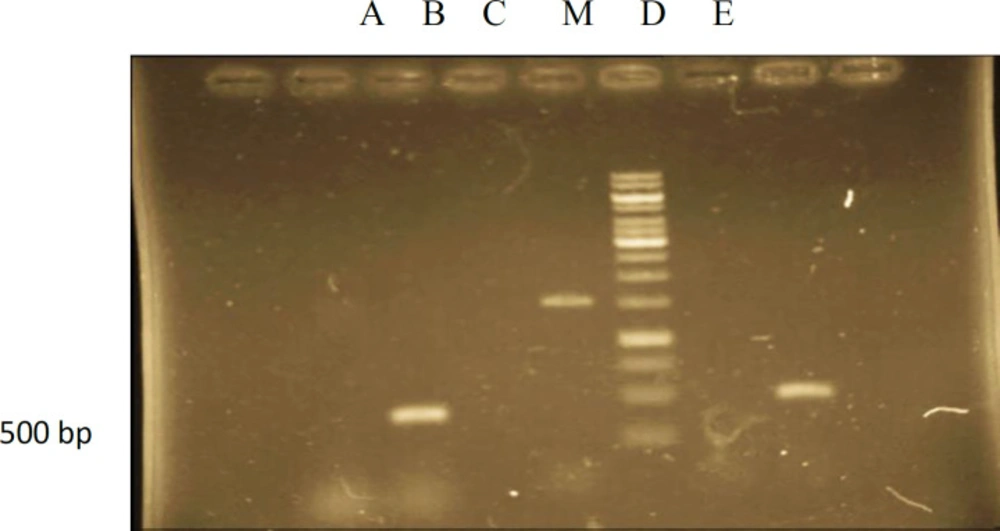
PCR analysis of rol B gene
To affirm the joining of T-DNA from the
A.rhizogenes into the hairy root genomic DNA, DNA from hairy roots were subjected to PCR examination. PCR was utilized to show that the T-DNA from the Ri plasmid of
A.rhizogenes was available in
A.komarovii genome and
rol B is one gene of the TL-DNA (T-DNA left arm) of Ri plasmid in
A.rhizogenes.
A.komarovii changed with the
rol B gene of
A. rhizogenes during the ceaseless sub-culturing. In this investigation, by utilizing DNAs from the hairy roots as template and non-transform roots as a control, PCR products opened up with
rol B forward and switch primers could be identified It was exhibited that every single changed root demonstrated the 500 bp
rol B gene and there was no
rol B gene found in control roots (
Figure 2). The biochemical and molecular characterization of the T-DNA genes from
A.rhizogenes is not completely comprehended and in a few investigations, proposing that new meristem development and consequent separation of changed plant cells might be directed through complex molecular systems (
19). To affirm the joining of T-DNA from the
A.rhizogenes into the hairy root genomic DNA, DNA from hairy roots were subjected to PCR examination. PCR was utilized to show that the T-DNA from the Ri plasmid of
A.rhizogenes was available in
A.komarovii genome and
rol B is one gene of the TL-DNA (T-DNA left arm) of Ri plasmid in
A.rhizogenes.
A.komarovii changed with the
rol B gene of
A. rhizogenes during the ceaseless sub-culturing. In this investigation, by using DNAs from hairy roots as template and non transform roots as a control, PCR products opened up with
rol B forward and the switch primers could be identified (
20). We performed DNA investigation, be aware of that end goal to affirm the hairy roots change. The TL district in plasmid T-DNA agropine sort strain
Agrobacterium rhizogenes ATCC15834 contain 18 operon perusing outlines including a few loci (root loci) (and the impact of TR and TL locales of
A. rhizogenes on development hairy roots (
21). Numerous different were additionally brought up that the
rol genes are actuated hairy roots and
rol B is an effective inducer of secondary metabolism in transgenic plants (
22). In this examination increment in the quantity of hairy roots the branches was practically logarithmic and we revealed high scopolamine creation by hairy root of
A.komarovii changed with the
rol B gene of
A. rhizogenes amid of persistent sub-culturing. Numerous different reports additionally brought up that the
rol genes are prompted hairy roots and
rol B is a capable inducer secondary metabolism in transgenic plants (
23). Hairy roots quickly developed with stable and relatively high substance in secondary metabolites and actuated by genetic change of
A.rhizogenes, were effectively utilized for the creation of secondary metabolites from medicinal or fragrant plants and genetic changed root cultures may deliver the secondary metabolites like that of the intact plants (
24).
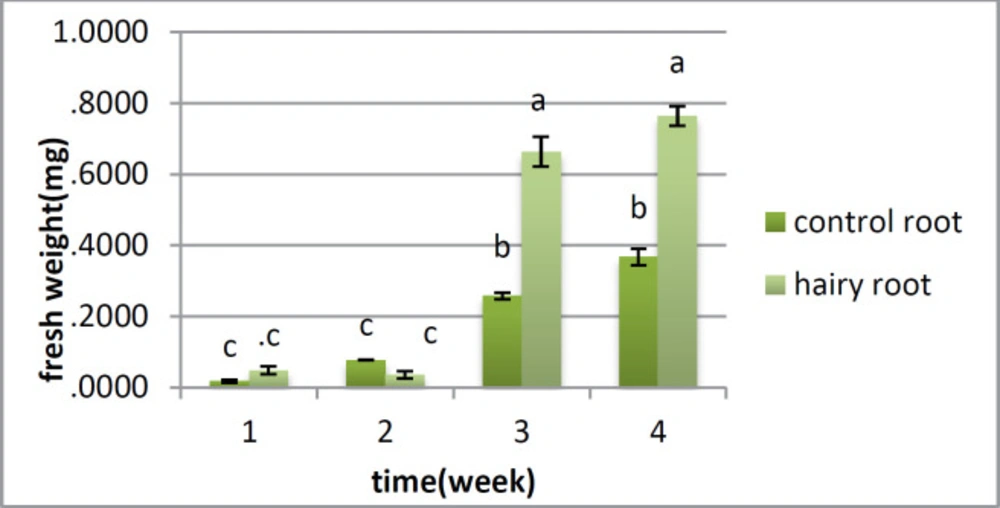
Analysis for fresh weight and dry weight
There is no distinction between the hairy root and control root in fresh weight in the first and second weeks; however the huge change was found in the third and fourth weeks, when the fresh weight of the roots was higher than that of the control roots (
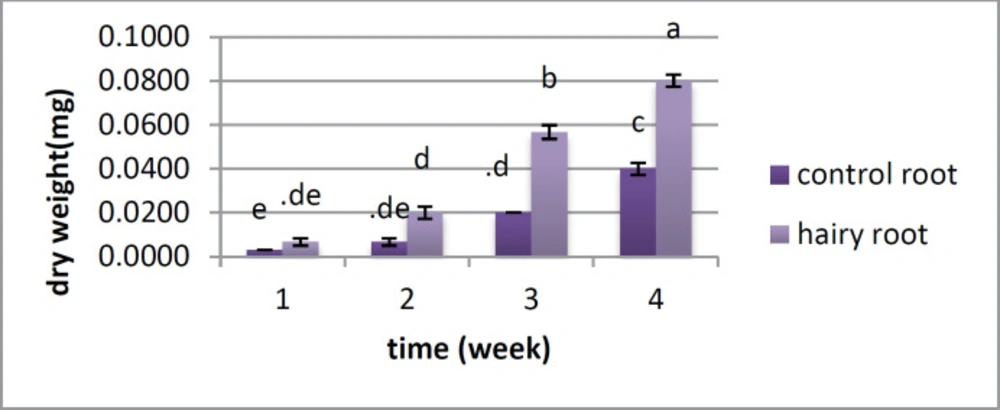
Figure 3). Dry weight showed a critical distinction between the two samples from the first week, and this distinction was observed over a month. In the first week, there was no huge distinction between them; however there was a contrast between the hairy roots and control roots from the second to the fifth week (
Figure 4).
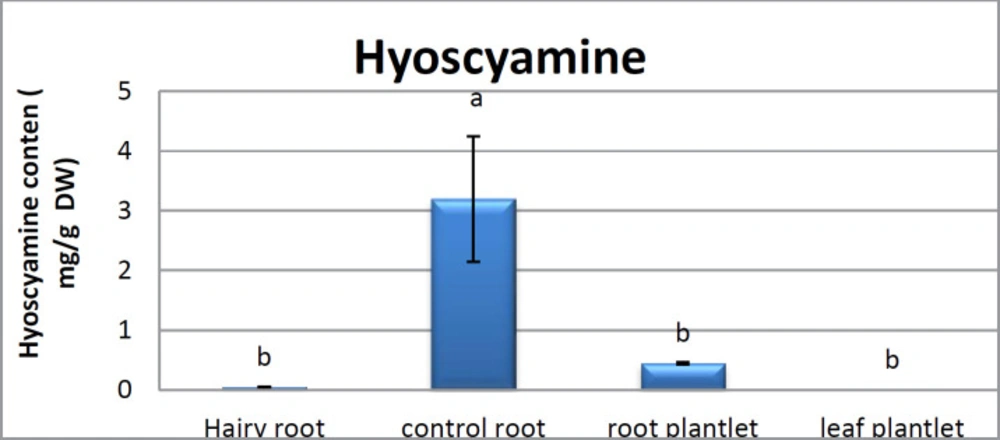
Scopolamine and hyoscyamine analysis
The hairy roots quickly developed with stable and relatively high substance in secondary metabolites, actuated by genetic change of
A.rhizogenes, were effectively used for the creation of secondary metabolites from medicinal or fragrant plants and the genetic changed root cultures may deliver the secondary metabolites like that of the intact plants (
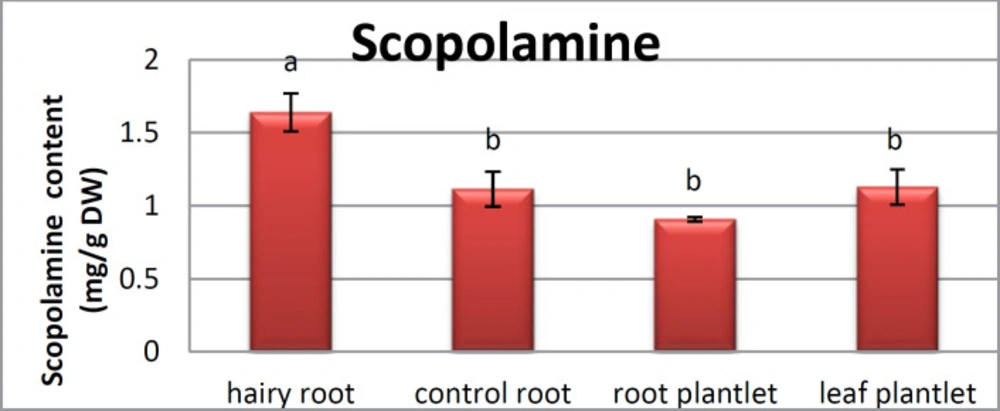
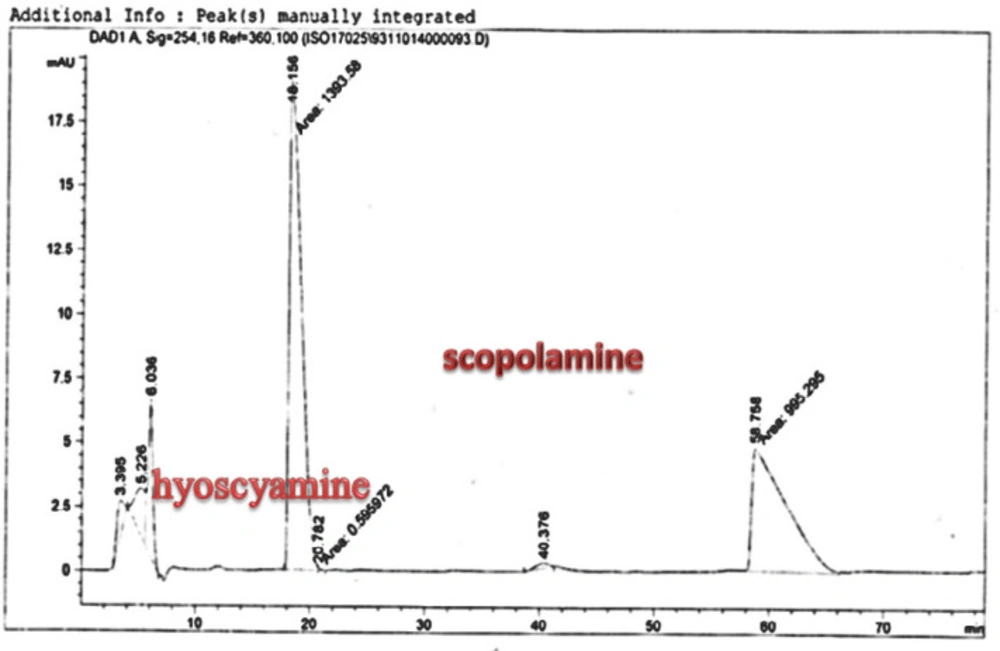
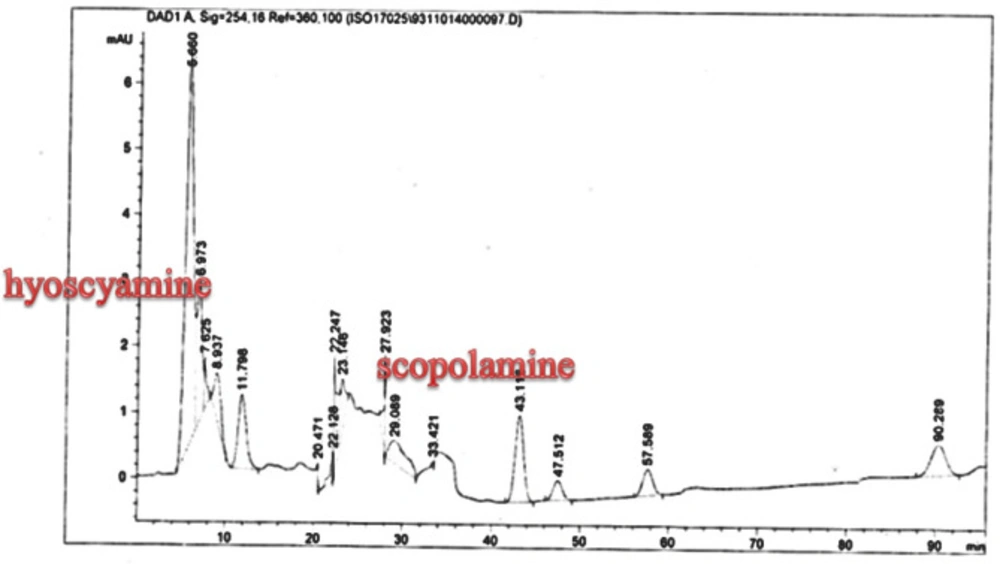
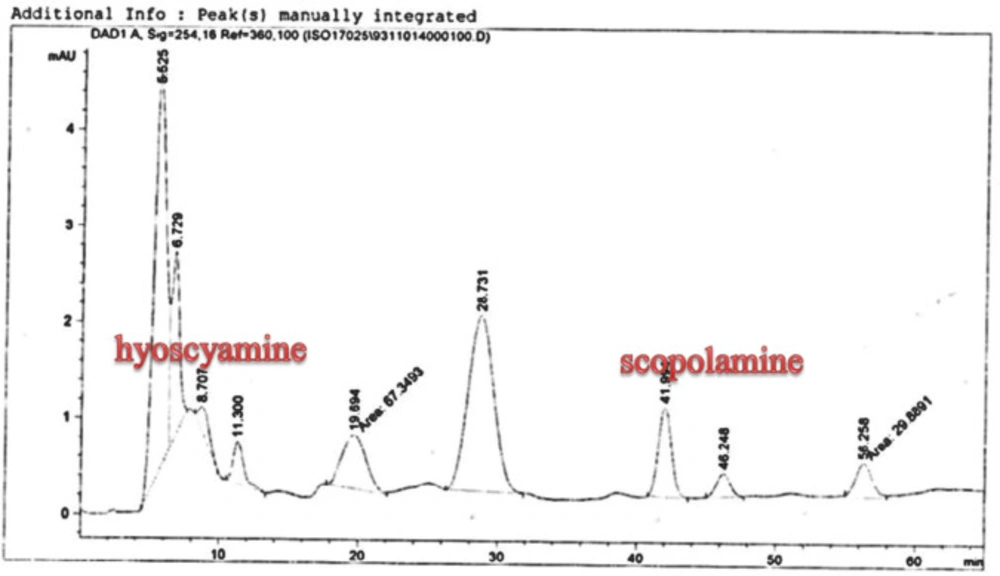
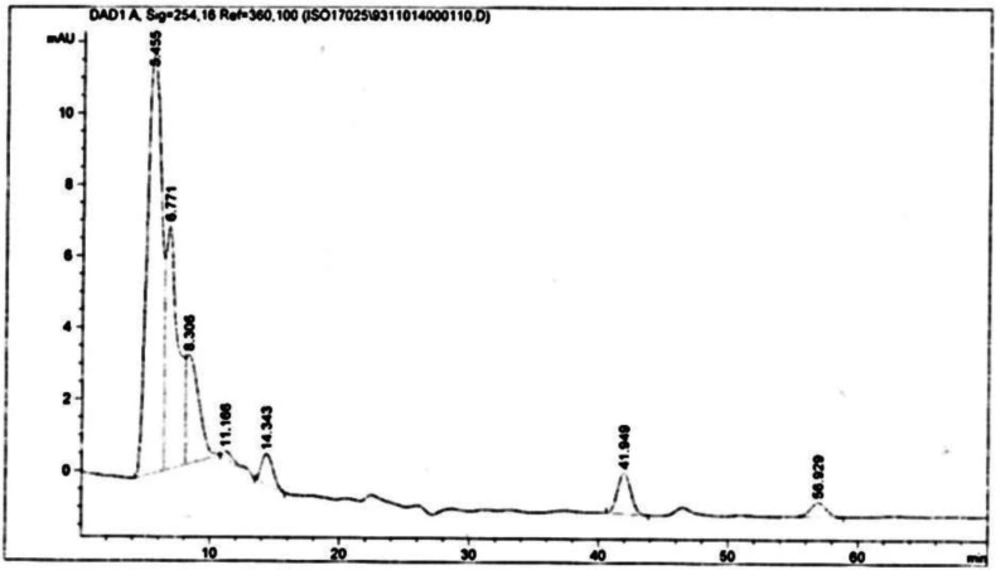
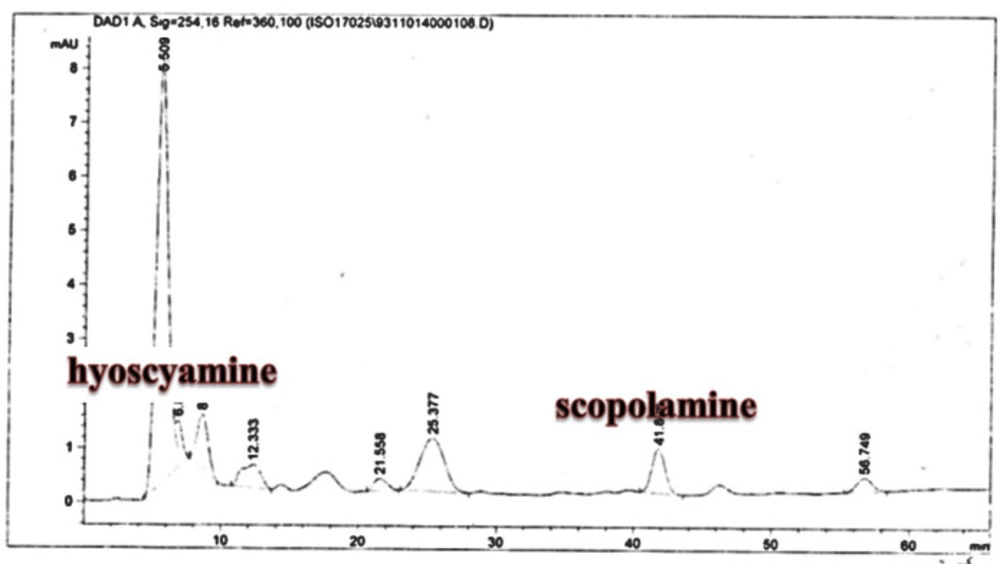
25). The substance of scopolamine and hyoscyamine were recognized in the hairy roots, non-transformed roots, roots, and leaves of the plantlet and both of these tropane alkaloids could be identified in all samples The maintenance time of the scopolamine substance of the hairy root culture was contrasted with the standard scopolamine and hyoscyamine (Sigma) In the HPLC profile, the RT value indicated the same points for both the standard and the concentration of the hairy root culture (
Figure7). The outcomes concerning the primary tropane alkaloids (scopolamine and hyoscyamine) from
Atropa komarovii plant tissue with hairy roots demonstrate higher measure of scopolamine than hyoscyamine in hairy roots (
Figure 5). From the outcomes concerning the fundamental oppositely, we found that the greatest measure of hyoscyamine was available in the control roots and on the other hand, scopolamine was the highest content compared to other treatments such as a leaves and roots of
Atropa komarovii plantlet (
Figure 5 and
6). Pathway-designing hairy root cultures are not just the perfect arrangement of recognize genetic capacities and in the past reports, transgenic hairy root cultures of
Atropa belladonna were stablished by over expressing different genes with the biosynthetic pathway of tropane alkaloids (
26). This result indicates that concurrence with information on scopolamine aggregation in hairy roots of
Atropa belladonna and shows that there are many elements participate in scale-up scopolamine in tissue culture revealing that expansion of scopolamine content in hairy roots of
A. belladonna by means of bioreactor (
27). We observed a considerable increment of scopolamine content in hairy roots and hyoscyamine was increased in nontransformed roots (
Figure 6,
8,
9,
10 and
11). Biosynthesis of secondary metabolite in changed roots is genetically controlled. In many reports in other genera of solanaceae transgene, invigorated scopolamine creation and its scopolamine synthesis appears to be metabolic regulation (
7). The nutritive factors on the production of two tropane alkaloids, scopolamine, and hyoscyamine in
Atropa belladonna hairy roots, the reserchers observed that the amount of hyoscyamine increased in hairy roots compared to plant leaves and roots. This results opposed to our studies and scopolamine content in the hairy roots were 10 times higher than in organs of intact plants (
Figure 6). Chashemi
et al. indicated that synthesis of hyoscyamine and scopolamine in
Atropa belladonna hairy roots increased than control roots and the hyoscyomaine is more than scopolamine in hairy roots and it was against of our reports (
22). Tropane alkaloid distribution in
Atropa baetica plants and they said hyoscyamine was more abundant, with the highest concentration in the main root followed by leaves and then scopolamine present in highest concentration in the main root (
26). In our researches, hyoscyamine in control root was higher than root plantlet and hairy roots and scopolamine were obsereved in all the samples although it has the highest amounts in hairy roots. Banerjee
et al. studied the expression tropane alkaloids in hairy root culture of
Atropa acuminate scopolamine that was higher in hairy root than hyoscyamine and the major tropane alkaloids was scopolamine and hyoscyamine in
Atropa acuminate (
24).