Protein-protein interaction (PPI) network analysis has a significant growth in cancer
studies to facilitate introducing early stage biomarkers (
23). In our study, the laryngeal cancer related proteins were analyzed via PPI
network construction, hub gene identification, module analysis, and functional enrichment
analysis of most significant modules. These stages were carried out for up-regulated
proteins and down-regulated ones in human laryngeal cancer tissue, separately. As it is
shown in
Tables 1 and
2, there are 275 changed expression proteins (including up and down regulated
proteins) related to the human tissue of laryngeal cancer. Data management and analysis is a
difficult process due to huge numbers of the collected proteins. Since PPI network analysis
is a powerful method in categorization and ranking of the candidate and related proteins for
a certain disease, here the up and down regulated networks are constructed separately (
Figures 1 and
2).
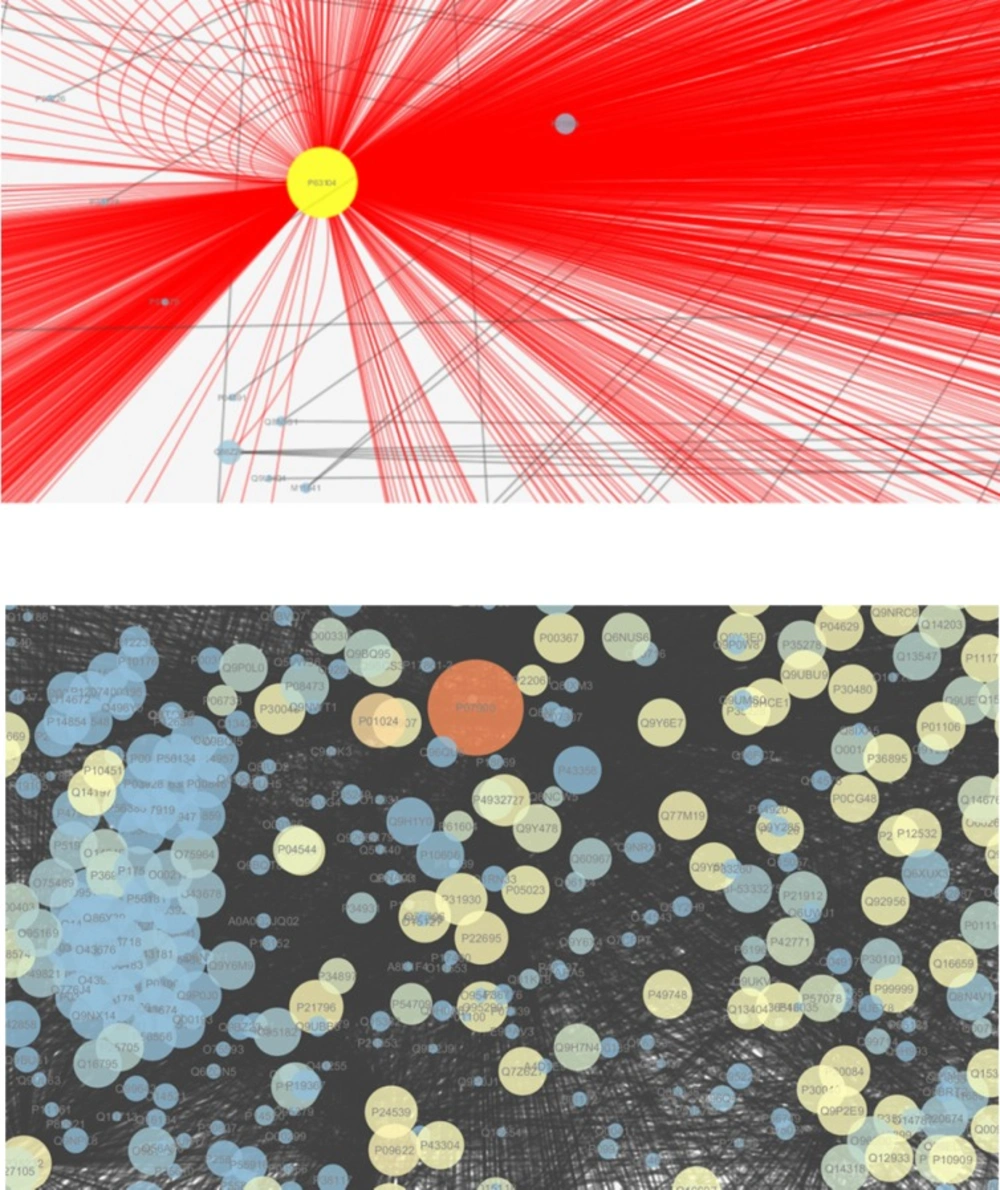
Topological analysis of the networks lead to rank of the nodes based on networks properties
(
18). By using two centrality indices including
degree and betweenness, totally 80 nodes are selected among 275 initial proteins as
important proteins (see
Tables 3 and
4). However, the number of 80 nodes can not be considered
as a suitable biomarker panel related to laryngeal cancer and more screening is required.
The hub-bottleneck nodes for the up and down regulated networks are shown in
Table 3. As it is shown in this Table there are 15 and 11
hub-bottlenecks for up and down regulated networks respectively. Module is a part of a
network including closed related proteins havig specific biological function (
20). Determined modules of network can provide
informative perspective about different roles of the nodes (
24). As it is shown in
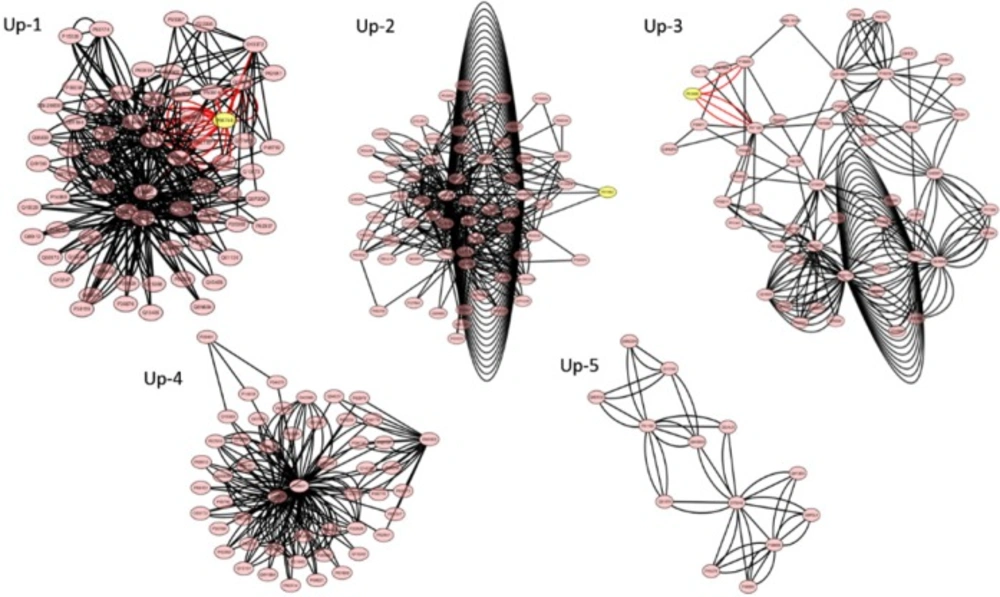
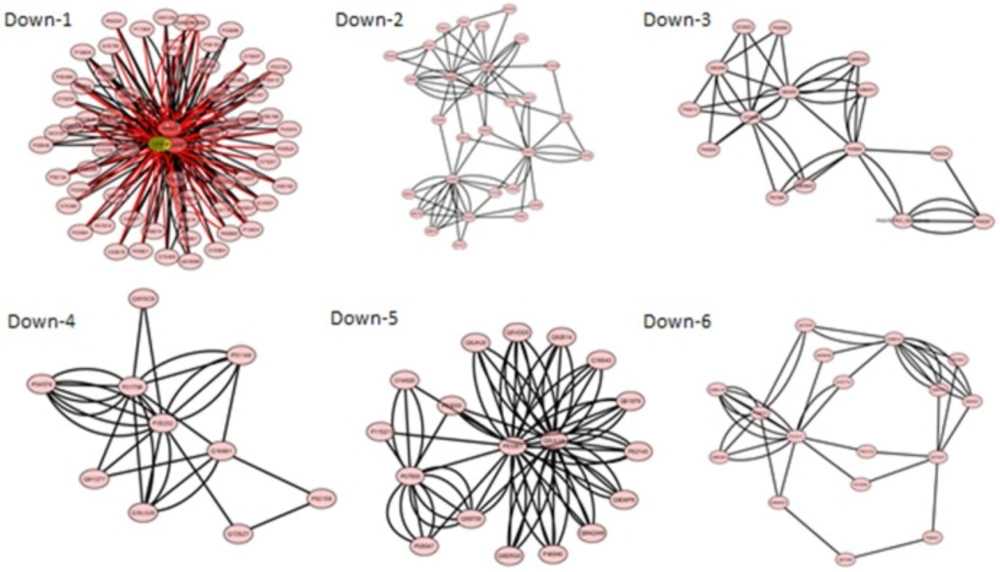
Figures 3 and 4 and
Table 5 there are 5 and 6 modules for the up and
down regulated networks respectively. Functional enrichment analysis for top score modules
indicated that RNA processing and splicing, mitotic cell cycle regulation and sugar
biosynthesis are affected by up-regulated modules while metabolic pathways and mitochondria
are the main affected subjects by down regulated modules (see
Table 6). The most significant pathways in four modules Up, 1-4 were RNA
processing, G1/S transition mitotic cell cycle, protein ubiquitination and RNA splicing. It
has been revealed overlapping between important pathways involved in the conversion of
pre-mRNA to mature mRNA. In previous studies, it shows that polymorphisms of mRNA processing
genes can be considered as risk factors for development of laryngeal cancer (
25). The most significant pathways in down regulated
modules (Down, 1-3) were respiratory electron transport chain, regulation of protein
insertion in to mitochondrial membrane involved in apoptotic signaling pathway, and NADH
dehydrogenase complex assembly. Proliferating cancer cells, such as laryngeal cancer,
preferentially use anaerobic glycolysis rather than oxidative phosphorylation for energy
production (
26). In one system biology study, the
glycolysis/gluconeogenesis pathway has been introduced as the most important pathway in
laryngeal cancer (
27). Then, the production of energy
from mitochondrial respiratory may shift to glycolysis in laryngeal cancer. To prove this
hypothesis and determine the energy supply sources of laryngeal cancer cells, more studies
are needed. Regulation of protein insertion into mitochondrial membrane involved in
apoptotic signaling pathway is the other important pathway in down regulated modules. One of
the mechanisms impaired cancer cells is apoptosis. Apoptosis can be activated through
several different signaling pathways, but a part of this mechanism is controlled in
mitochondrial membrane through insertion apoptotic proteins (
28). According to these results, in laryngeal cancer, apoptotic mechanism may
disturb through the impairment of transporter proteins which transform apoptotic proteins
into mitochondria. According the results of
Table 5,
the scattering of hubs in up-modules was more than down ones. Interestingly, the finding
indicate that the seeds and hubs in up-modules have the similar functions with each other
that are associated with regulation of cell cycle (
29,
30). Among 26 hub-bottleneck nodes 12
proteins (8 up-regulated and 4 down-regulated proteins) are distributed in 8 modules (see
Table 5). These proteins are tabulated in
supplementary Table S1 and are ranked based on amounts of degree value. Here two suggestions
are feasible: first investigation about expression changes of these 12 genes in the field
and the second idea is selection of the top up and down regulated genes for more
examinations. We choose cutoff 1200 for degree and therefore YWHAZ and PPP2R1A as the top
two up-regulated genes and also HSP90AA1 and CALM3 as the top two down-regulated genes are
introduced as human laryngeal cancer. YWHAZ gene with the highest degree and BC scores
encodes 14-3-3 protein zeta/delta that has an essential role in tumor cell proliferation
(
31) through the regulation of multiple cellular
processes, such as cell cycle control, anti-apoptosis, signal transduction, inflammation,
and cell adhesion/motility (
32). YWHAZ has been
introduced as candidate proto-oncogene in head and neck squamous cell carcinoma whose
reduced expression causes lower level of DNA synthesis rates (
33). 14-3-3 proteins could be a key regulatory components in many
processes that are crucial for development of cancers (
34) such as laryngeal cancer (
8). PPP2R1A
gene encodes one subunit of protein phosphatase 2. This protein phosphatase is involved in
control of cell growth and cell division processes. The role of this subunit in integrity of
enzyme is highlighted. Therefore, it is expected that PPP2R1A plays a crucial regulatory
role in cell proliferation in cancer cell line(
35).
HSP90AA1 and CALM3 were found as two top ranked genes in the down-regulated PPI network.
These proteins belong to family of proteins which involved in the regulation of specific
target proteins in cell cycle control and programmed cell death (
36,
37). On the other hand, CALMs
in addition to cell cycle, related to centrosome cycle and deregulation of this protein can
be the origin of chromosomal instability in cancer (
38). Interestingly, all determined possible biomarkers are related to the cell
cycle process.