



1. Background

The chemic structures of A, lamotrigine; and B, phenobarbital
2. Methods
2.1. Materials
2.2. Apparatus and Procedure
2.2.1. Fluorescence Study
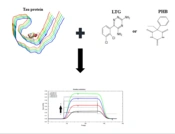
2.2.2. Tau Protein Immobilization Procedure
2.2.3. Interactions of Lamotrigine and Phenobarbital with Immobilized Tau Protein
2.2.4. Molecular Docking Studies
3. Results and Discussion
3.1. Tau Protein Fluorescence Quenching
/B, phenobarbital (PHB) interactions as determined by intrinsic fluorescence emission intensity in the presence of additional concentrations of LTG (0, 4, 19.5, 39, 58.5, and 78 μM) and PHB (0, 4, 8.5, 13, 17 and 21.5 μM) in phosphate buffer (pH 7.4) at three different temperatures, 298, 310, and 316 K")
Tau protein and A, lamotrigine (LTG)/B, phenobarbital (PHB) interactions as determined by intrinsic fluorescence emission intensity in the presence of additional concentrations of LTG (0, 4, 19.5, 39, 58.5, and 78 μM) and PHB (0, 4, 8.5, 13, 17 and 21.5 μM) in phosphate buffer (pH 7.4) at three different temperatures, 298, 310, and 316 K
 and B, phenobarbital (PHB) at three different temperatures (298, 310, and 316 K) and various concentrations and double logarithmic curves for tau protein in the presence of different concentrations of C, LTG; and D, PHB at three various temperatures (298, 310, and 316 K)")
Fluorescence quenching Stern-Volmer plot of tau protein with increasing concentrations of A, lamotrigine (LTG) and B, phenobarbital (PHB) at three different temperatures (298, 310, and 316 K) and various concentrations and double logarithmic curves for tau protein in the presence of different concentrations of C, LTG; and D, PHB at three various temperatures (298, 310, and 316 K)
| Sample and Temperature (K) | Stern-Volmer Quenching Constants (× 104 M-1) | Bimolecular Quenching Constants (× 1012 M-1.s-1) | R2 |
|---|---|---|---|
| Lamotrigine-tau | |||
| 298 | 2.05 | 2.05 | 0.9844 |
| 310 | 1.52 | 1.52 | 0.9841 |
| 316 | 1.15 | 1.15 | 0.9926 |
| Phenobarbital-tau | |||
| 298 | 4.59 | 4.59 | 0.9866 |
| 310 | 3.42 | 3.42 | 0.9881 |
| 316 | 2.12 | 2.12 | 0.9301 |
3.2. Binding Constants and Binding Sites
| System and Temperature (K) | Binding Constants (× 104 M-1) | Stoichiometry of Binding | ∆H (kJ.mol-1) | ∆S (kJ.K-1.mol-1) | ΔG (kJ.mol-1) | R2 |
|---|---|---|---|---|---|---|
| Lamotrigine-tau | -46.76 | -0.080 | ||||
| 298 | 0.938 | 0.92 | -22.92 | 0.9833 | ||
| 310 | 0.573 | 0.90 | -21.96 | 0.9711 | ||
| 316 | 0.300 | 0.86 | -21.48 | 0.9964 | ||
| Phenobarbital-tau | -36.70 | -0.035 | ||||
| 298 | 3.415 | 0.97 | -26.27 | 0.9735 | ||
| 310 | 2.349 | 0.96 | -25.85 | 0.988 | ||
| 316 | 1.191 | 0.95 | -25.64 | 0.9224 |
3.3. Thermodynamic Parameters Analysis
 and lamotrigine (LTG) with tau protein obtained from fluorescence data analysis; B, the plots for PHB to tau protein; and C, LTG to tau protein obtained from surface plasmon resonance analyses")
A, van’t Hoff plots for the interactions of phenobarbital (PHB) and lamotrigine (LTG) with tau protein obtained from fluorescence data analysis; B, the plots for PHB to tau protein; and C, LTG to tau protein obtained from surface plasmon resonance analyses
3.4. Surface Plasmon Resonance Parameter Analyses
 at three different temperatures 298 K, 310 K, 316 K")
Sensorgram of lamotrigine-tau protein at different concentrations (25, 50, 100, and 150 μM) at three different temperatures 298 K, 310 K, 316 K
| System and Temperature (K) | Association Rate Constants (M-1.s-1) | Dissociation Rate Constants (s-1) | Equilibrium Constants (M) | ∆H (kJ.mol-1) | ∆S (kJ.K-1.mol-1) | ΔG (kJ.mol-1) |
|---|---|---|---|---|---|---|
| Lamotrigine-tau | 554.77 | 1.616 | ||||
| 298 | 1.20 × 102 | 3.37 × 10-11 | 2.80 × 10-13 | 73.208 | ||
| 310 | 1.44 × 104 | 2.18 × 10-6 | 1.51 × 10-10 | 53.816 | ||
| 316 | 1.41 × 105 | 2.61 × 10-2 | 1.85 × 10-7 | 44.12 | ||
| Phenobarbital-tau | -815.40 | -279.11 | ||||
| 298 | 7.91 × 101 | 3.52 × 10-2 | 4.45 × 10-4 | 82359 | ||
| 310 | 2.92 × 105 | 3.71 × 10-2 | 1.27 × 10-7 | 85708 | ||
| 316 | 3.25 × 106 | 3.06 × 10-6 | 9.42 × 10-13 | 87383 |
3.5. Molecular Docking Study
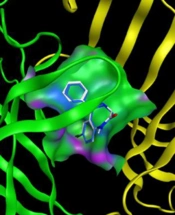
 to tau protein, A, the binding site; B and C, detailed illustration of LTG-tau protein complex, and also D, the binding mode of phenobarbital (PHB) with tau protein; E and F, detailed illustration of PHB-tau protein complex in S1 pockets.")
Molecular docking model of the binding of lamotrigine (LTG) to tau protein, A, the binding site; B and C, detailed illustration of LTG-tau protein complex, and also D, the binding mode of phenobarbital (PHB) with tau protein; E and F, detailed illustration of PHB-tau protein complex in S1 pockets.