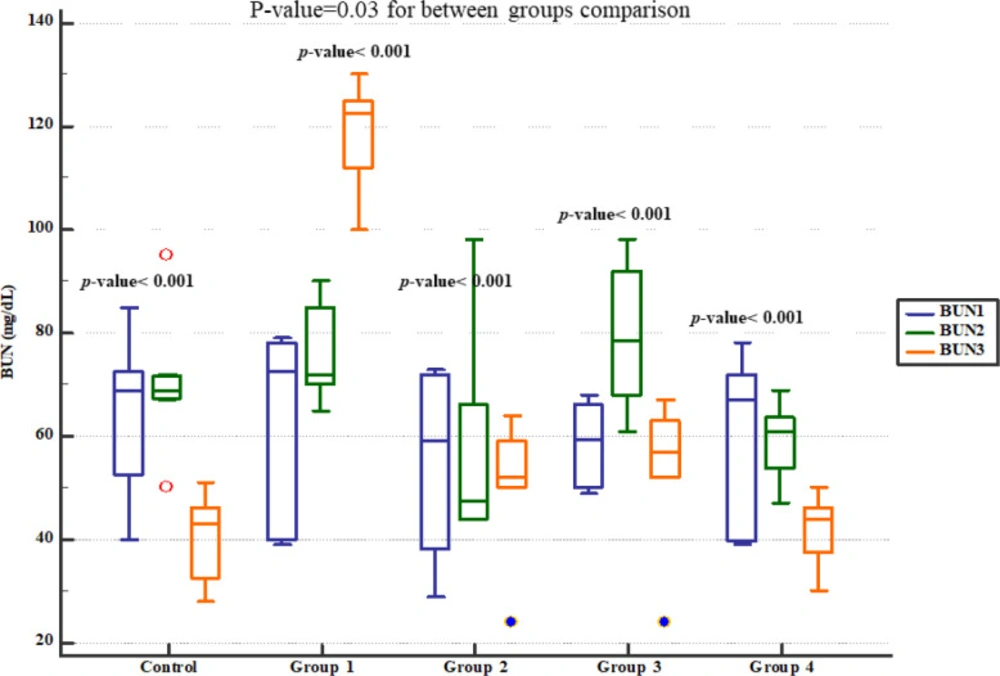
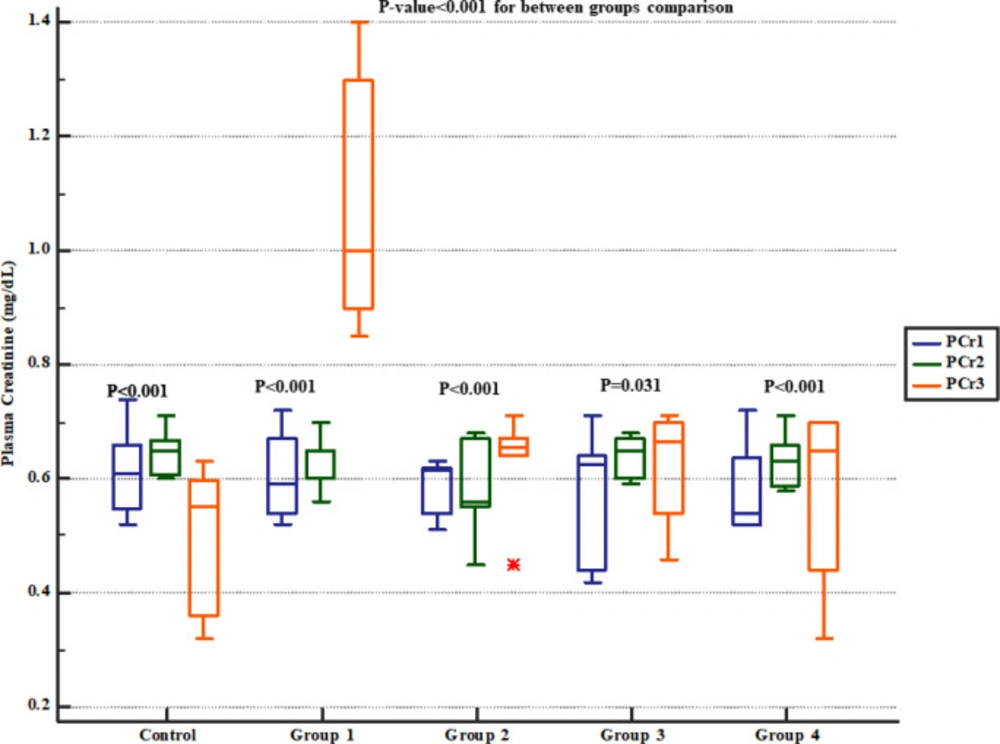
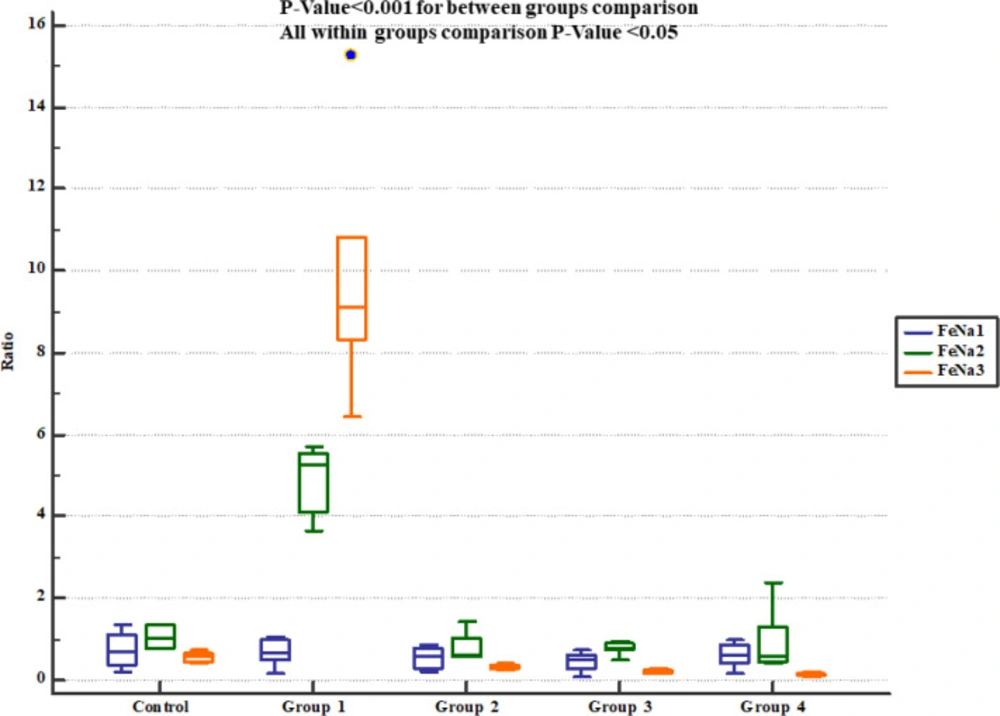
However, there were significant and dose-dependent reductions in serum BUN, creatinine, cholesterol and triglyceride, urinary Na
+ excretion, apoptosis rate, and inflammatory cells infiltration in renal tissues. Other studies have also evaluated protective effects of various agents against Gm-induced AKI and reported similar findings as increased serum BUN and creatinine levels in animal models (
5,
8,
32). Increased levels of BUN and creatinine in the context of Gm-induced renal impairment have been correlated with prominent aberrations in renal function and structure (
33). In line, Gm administration for 8 days reduced creatinine clearance and renal blood flow in previous reports (
32,
34). The rats exposed to Gm have also shown dose-dependent decline in glomerular filtration rate (
29,
35). In support, Troxerutin,a plant flavonoid, was able to correct alternations in urinary protein and albumin levels, as well as serum creatinine and BUN in part by augmenting glomerular filtration rate (
29). Although elevations in serum BUN and creatinine indicate renal functional impairment, these traditional markers may not be adequately sensitive to reveal renal toxicity in early phases (
36). Some novel markers including urinary neutrophil gelatinase associated protein (NGAL), β
2-microglobulin, albumin, clusterin, glucose excretion, histological findings, urinary level of kidney injury molecule-1 (KIM-1) and alpha glutathione S-transferase have been suggested to develop a multisystem model for assessing drug induced renal injury (
36-
38). In mice and HEK293 cells, Gm exposure increased urinary levels of KIM-1, NGAL, and Cys-C activities (
8,
29). In addition, as early as four days post Gm-exposition, increased expression of four miRNAs (miR-138-5p miR-1971, miR-218-1-3p, and miR-489) predicted Gm-induced toxicity before elevation of serum BUN and creatinine (
39). With advancements in diagnostic methods, novel biomarkers have been proposed to be used as alternatives to serum BUN and creatinine to early detect drug-induced AKI. In current study, pre-treatment with either 800, 1600 or 2400 mg/Kg of
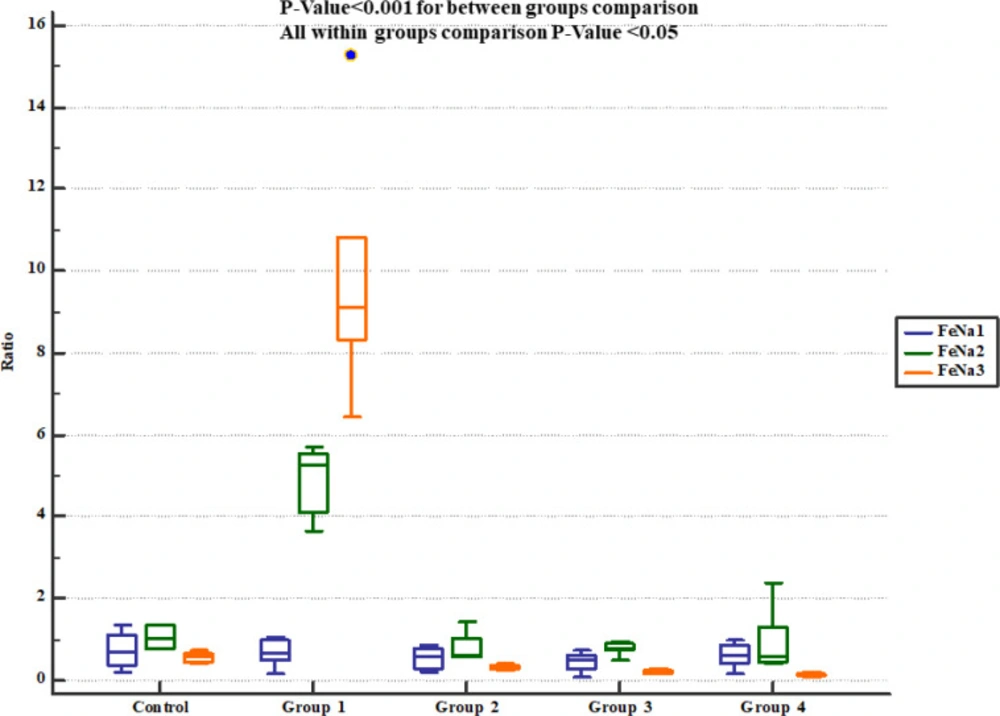
Descurainia sophia significantly decreased urine Na
+ concentration and FeNa
+ at all the studied doses. In accordance, Gm treatment exerted similar patterns in Na
+ levels in previous studies (
34,
40). Also previous experiments showed that Gm treatment significantly increased FeNa
+ level (
41,
42). Alternations in electrolytes distribution in Gm-induced AKI can be related to the depressed activity of Na
+, K
+-ATPase pump in proximal renal tubules correlating with higher Na
+ excretion (
43). Nevertheless, Sugarman et al described that Gm had no significant impact on the activity of this transporter in renal medullary or cortical cells (
35). Gm-induced accumulation and release of Na
+ and K
+ respectively from renal proximal tubular cells can also contribute to the disturbed electrolyte turnover in Gm-induced nephrotoxicity (
44). In fact, the concentration dysfunction in tubular cells, rather than defects in sodium excretion disturbance, has been suggested as the main cause of disturbed Na
+ distribution induced by Gm (
35). According to studies, Gm damages tubular tissue in kidneys resulting in functional disturbances of tubular cells in regulating the balance of electrolytes and other molecules (
45). Trace elements perform important biological roles in the body including participation as structural elements in enzymes, proteins, and hormones (
46), and using agents protecting electrolyte disturbance can result in healthy renal and other organs functions. Pre-treatment with either 800, 1600, or 2400 mg/kg
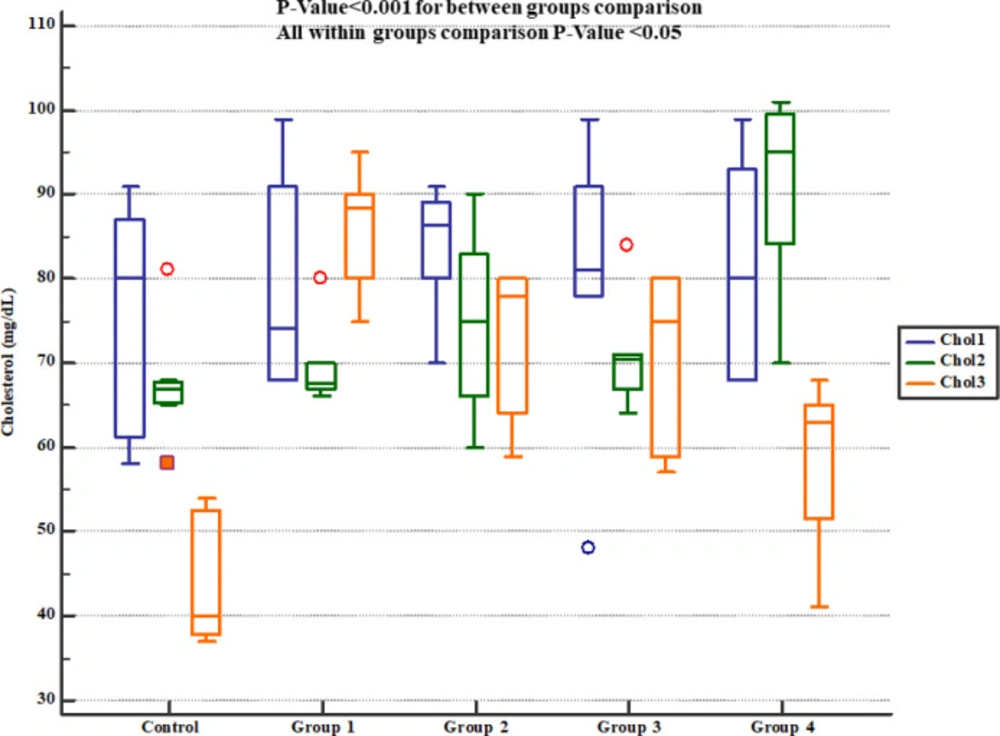
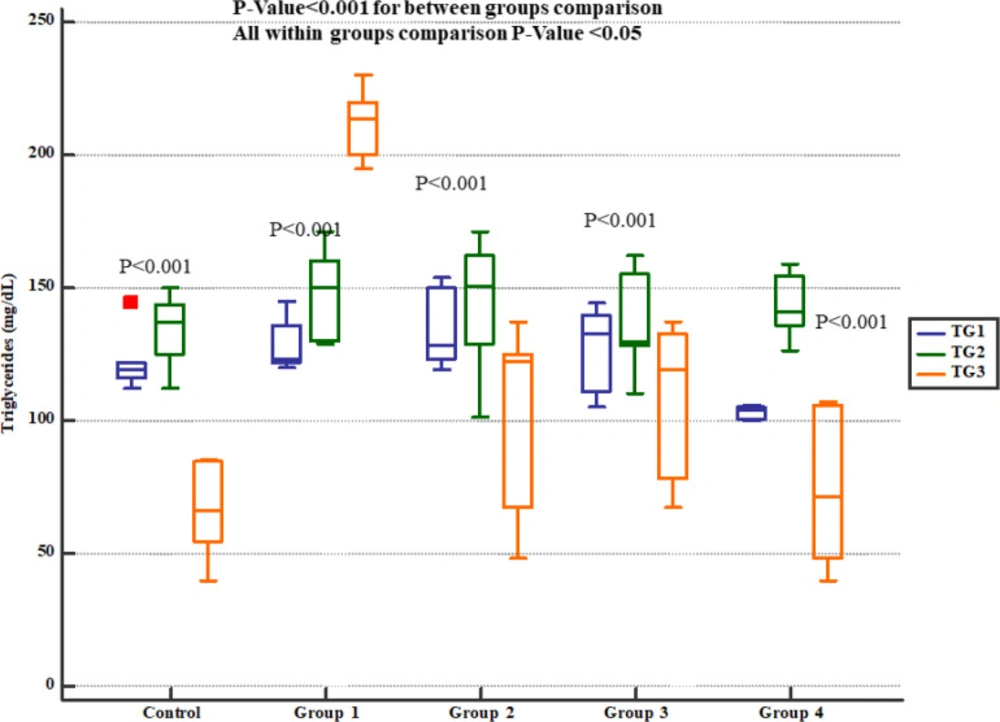
Descurainia sophia prevented increments in serum cholesterol and triglycerides levels in Wistar rats. All the doses were significantly efficient; however, the most potent effects were noticed in the dose of 2400 mg/Kg. In line with this, Anandan et al in their study also noted that hesperidin effectively prevented Gm-induced elevations of serum cholesterol, free fatty acids, and triglycerides in rats (
30). It has been shown that Gm can damage lysosomes resulting in high levels of membrane lipids (sphingomyelin and sphingosine) in serum (
47). However, studies evaluating the effects of Gm on lipid profile and its interaction with renoprotective pathways are scare and more studies are needed in this area. Moreover, the results of this study showed that treatment with
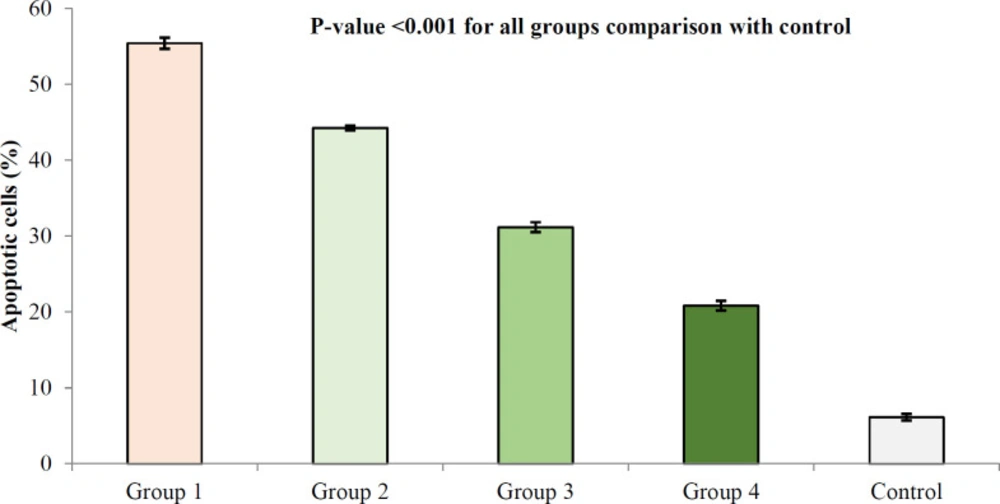
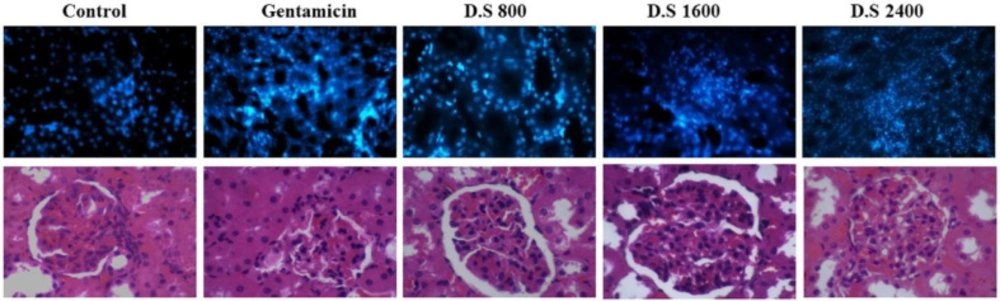
Descurainia sophia pre-administration dose-dependently protected renal cells from Gm induced apoptosis. This observation was in line with a previous report indicating higher apoptosis rate in Gm-induced renal cells (
29). Accordingly, after 14 days of administration, Gm induced apoptosis in renal tubular cells in rats (
5). In addition to
in-vivo models, Gm also induced dose-dependent apoptosis in HEK293 (
1,
8) and HK-2 (
1) kidney cell lines. Gm was shown to activate Bax; a proapoptotic member of Bcl-2 family, which subsequently triggers caspase-3 dependent cellular death (
48). Increased expression of Fas ligand (CD95) in renal cells can also contribute to Gm-induced apoptosis (
32). In accordance with our results,
Descurainia sophia extract reduced apoptosis rate in cardiomyocytes of rat model of cardiac disease which was concomitant with depressed levels of Bax and caspase 3 (
22). In A549 tumor cells,
Descurainia sophia seed extract induced the expression of death receptors 4 (DR4) and 5 (DR5) through activating endoplasmic reticulum stress response pathway (
49). In another study, dapagliflozin inhibited Gm-induced renal tubular cells apoptosis in part by increasing miR-21 and decreasing miR-181a expressions (
5). Overall, the exact anti-apoptotic mechanisms of
Descurainia sophia in Gm-induced renal injury are to be elucidated.
Finally, Gm treatment for 7 consecutive days resulted in renal cellular damage and infiltration of inflammatory cells in kidney tissue in the rats receiving saline. Pre-treatment with
Descurainia sophia significantly reduced cellular injuries and recruitment of inflammatory cells to kidney tissue (
Figure 9). An interstitial nephritis may also accompany the tubular damage by Gm (
53-57). A lymphoproliferative phenomenon was also observed in renal tissue of Gm-exposed rats in a study by Oliveira et al (
50). In other studies, histological characteristics have been noted in Gm-treated kidneys as degenerative changes (
32,
51) and vaculation (
5) in tubular epithelial cells (
5,
36) and glomeruli (
30), proliferative glomerulonephritis (
32), tubular necrosis (
30,
33,
35,
40,
51,
52), inter-tubular bleeds (
5), vascular inflammation (
5), tubular hyaline casts formation (
5,
35,
37,
40) and dilation of renal tubules (
5,
37). Gm nephrotoxicity has been associated with ultrastructural changes in basal membrane of glomeruli and renal tubules as well (
53). Gm is filtrated through glomeruli, and during this process, binds to negatively charged structures triggering membrane permeability dysfunction and finally necrosis of tubular cells (
54). The essential role of tubular necrosis in Gm-induced renal toxicity is supported by the reversal of renal function after inhibition of tubular necrosis by pre-administration with dipyridamole (
33). In addition, dapagliflozin restored structural and functional capacities of kidneys in the rats treated with Gm through inhibiting renal tubular cells apoptosis (
5). Vitamin E (α-tocopherol) also corrected functional deficiencies of Gm-exposed kidneys in part through reversing structural changes in glomerular basal membrane and tubules (
53). Accordingly, as the role of inflammatory cells in histopathological events in Gm-induced renal injury has been indicated, the administration of anti-inflammatory agents such as
Descurainia sophia can be beneficial.
Gm-induced nephrotoxicity is mediated through a complex multifactorial phenomenon with various pathways participating in this process. Among the most important toxic effects of Gm is inducing oxidative stress by overproducing oxidative agents such as reactive oxygen species (ROS) and depressing anti-oxidative capacities in renal cells. Multiple reports have verified the role of oxidative stress in adverse renal effects of Gm in rats (
5,
8,
34,
50,
53). Anti-oxidant enzymes and mediators including super oxide dismutase (SOD), glutathione peroxidase, heme oxygenase -1, catalase and glutathione have been depressed in kidneys of Gm-treated rats (
6,
29,
30,
32). The rats treated with Gm have shown higher levels of malondialdehyde (MDA), a marker of lipid peroxidation, in renal tissue (
29,
32,
50). In fact many protective agents have exerted their effects at least in part by reducing oxidative markers and augmenting anti-oxidative mechanisms (
6,
8,
29,
34,
48,
51,
53). In line, the anti-oxidative effects of
Descurainia sophia have been established in multiple reports (
16-
18). Propagation of oxidants within renal tissue leads to structural changes in proteins, lipids, DNA, as well as functional exhaustion of critical organelles such as mitochondria (
53). Subsequently, these molecular events can trigger adverse pathological outcomes and tubular necrosis as one of the main underlying causes of Gm-induced nephrotoxicity (
55). Therefore, using anti-oxidative agents such as
Descurainia sophia can be helpful to prevent adverse effects of Gm on kidneys.
Gm also recruits inflammatory mediators to promote its nephrotoxic effects. This is supported by studies that showed beneficial effects of anti-inflammatory agents in preventing Gm-induced AKI (
48,
51). Elevated levels of inflammatory cytokines such as interleukin-6 (IL-6), IL-10, and tumor necrosis factor-α (TNF-α) have been described in renal tissue of models of Gm-induced AKI (
8,
29,
33). On the other hand,
Descurainia sophia seeds extract decreased the expression of IL-4, a major marker of type 2 helper T response and inflammation, in mouse model of asthma (
21). In mouse model of asthma,
Descurainia sophia seeds extract also downregulated VEGFa (
21). Other protective mechanisms by
Descurainia sophia may be exerted by encountering the processes initiated by Gm. For example, the plant extract through its active ingredients can modulate signaling pathways such as endoplasmic reticulum stress (i.e. unfolded protein response) (
49) and PI3k/Akt/mTOR pathways (
22). Helveticoside; an active ingredient of
Descurainia sophia was shown to role as a potent regulator of intracellular signaling pathways and gene expression (
56). Finally, ethanolic extract of
Descurainia sophia seeds suppressed the activity of cytochrome P450 isoforms (i.e. CYP1A2, CYP2C9, and CYP2C19) which are essential enzymes participating in metabolization of drugs (
57-
59). Other underlying mechanisms by which
Descurainia sophia extract and its ingredient suppress adverse effects of Gm on renal tissue are yet to be identified.