Cortical dis excitation developed absence seizures
Developmental alternations in the cortical excitation and/or inhibition are involved in the pathophysiology of some neurodevelopmental disorders (
24,
25). Cortical disinhibition has been indicated in the pathogenesis of Parkinson’s disease (
26). Cortical enhancement of excitability has a key role in the pathophysiology mechanism of Alzheimer’s disease. Further, cortical excitability might lead to cortical dysfunction in the primary lateral sclerosis as well as amyotrophic lateral sclerosis (
27,
28). It has been reported that the fluctuation of cortical excitation underlies hemiplegic attacks (
29). High resting, active motor threshold and small evoked potentials in patients with Sydenham’s chorea emphasized the low excitability of corticospinal output (
30).
Epilepsy as a neurodevelopment disorder has been investigated for cortical anomalies.
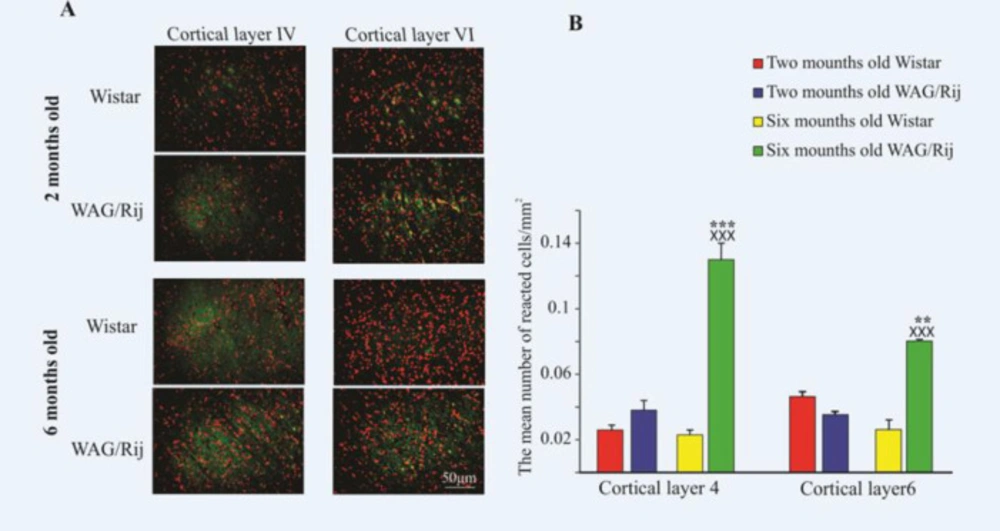
We evaluated the changes of the GluR1 subunit in the somatosensory cortex during the development of absence epilepsy.
WAG/Rij rats display SWDs in adulthood and are considered as a valid genetic model of gene-linked absence epilepsy (
9). Our findings indicated that the GluR1 subunit in the somatosensory cortex, in both IV and VI cortical layers, has been highly expressed in the adult epileptic rats. On the contrary, it has been down-regulated in the adult healthy Wistar rats.
GluR1 as a Ca
2+ permeable subunit of AMPARs is important for normal synaptic function after birth. In early developmental stages, many synapses contain GluR1, start to change into GluR2 (Ca
2+ impermeable subunit of AMPARs) during the age-dependent development of cortical layers (
31,
32).
Abnormalities of structure and function of synapses during the development of CNS are involved in the pathogenesis of absence seizures (
25). Prior studies have reported high expression of AMPA receptors in the hippocampus and cerebral cortex in the epileptic people and rats (
33-
35). In the hippocampus of epileptic people, the density of AMPA GluR1 subunit proteins has been increased (
36). GluR2 markedly has been down-regulated in the neurons that were subject to epileptic seizures (
37). Further investigations documented that enhancement of hippocampal GluR1 followed by some neuropathological conditions such as inflammation (
38) or impaired autophagy, reduced GluR1 degradation and increased seizure susceptibility in the adult rats (
39,
40). Functional and single-cell transcript analyses have shown that prolonged receptor responses in the hippocampal astrocytes of epileptic patients, resulted from high expression of GluR1 in this region (
41-
43).
These data suggest that aberrant patterns of physiological activities influence the composition of AMPA receptor subunits in a region-specific and/or cell-type-specific manner.
Synaptic activity is needed for synaptic delivery AMPA receptors containing the GluR1 subunit, whereas GluR2 is constitutively inserted into synapses (
44). It is previously was shown that the expression of the stargazin, a critical regulators of the trafficking and function of AMPA receptors, increased in the somatosensory cortex of genetic absence epilepsy rat from Strasbourg (GAERS) (
45); and its elevation is accompanied with an increase in AMPA receptor proteins (GluR1 and GluR2) in the somatosensory cortex of adult epileptic GAERS (
46). It seems that AMPA receptor subunits in different neuronal populations are subject to distinct regulatory processes in response to intense physiological activity.
Ictogenic neurons of deep layers of somatosensory cortex display distinctive hyperactivity that leads to these data suggesting that aberrant patterns of physiological activities influence the composition of AMPA receptor subunits in a region-specific and/or cell-type-specific manner.
Synaptic activity is needed for synaptic delivery AMPA receptors containing the GluR1 subunit, whereas GluR2 is constitutively inserted into synapses (
44). It is previously was shown that the expression of the stargazin, a critical regulator of the trafficking and function of AMPA receptors, increased in the somatosensory cortex of genetic absence epilepsy rat from Strasbourg (GAERS) (
45); and its elevation is accompanied with an increase in AMPA receptor proteins (GluR1 and GluR2) in the somatosensory cortex of adult epileptic GAERS (
46). It seems that AMPA receptor subunits in different neuronal populations are subject to distinct regulatory processes in response to intense physiological activity.
Ictogenic neurons of deep layers of somatosensory cortex display distinctive hyperactivity that leads to the firing of distant cortical and thalamic cells during the epileptic discharges (
5,
47 and
48). Based on these data we conclude that aberrant cortical expression of GluR1 receptors in epileptic rats may contribute to the development of SWDs.
AMPARs modulatory effects on the absence seizures
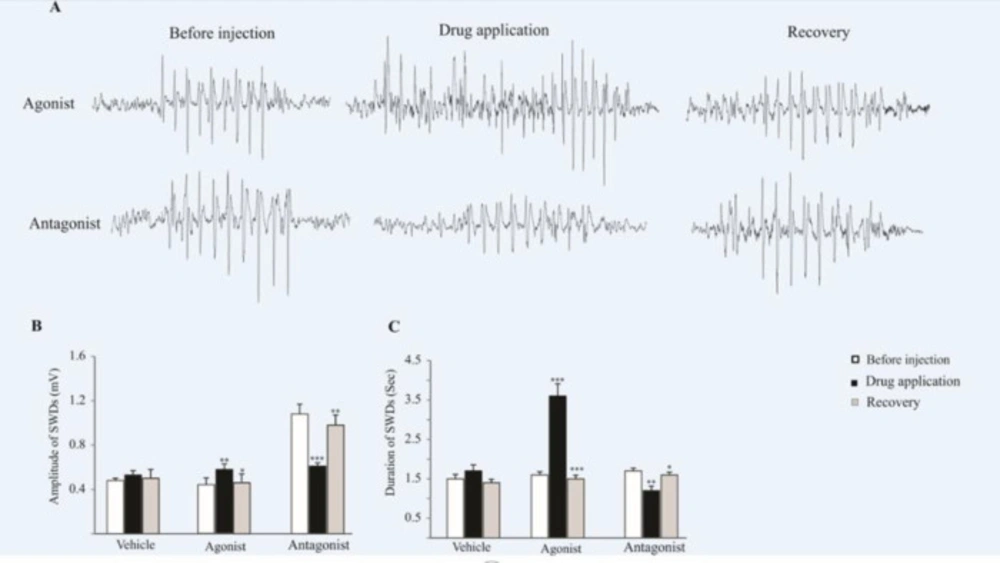
We assessed the effect of pharmacological modulation of AMPARs on the characteristic of bioelectrical brain activities in the somatosensory cortex of epileptic rats and showed that cortical activation of AMPARs exacerbated the absence seizures in the epileptic WAG/Rij rats.
Various pathophysiological mechanisms are involved in the abnormal neuronal discharges in epilepsy. Regardless of the primary cause, abnormal glutamate (excitatory) neurotransmission, affecting on metabotropic and ionotropic receptors, is was thought to be critical for seizure generation and the epileptic state (
21,
49 and
50).
Throughout the CNS, fast synaptic excitation within and between brain regions related to epilepsy is mediated mostly by AMPA and NMDA receptor localized to the postsynaptic membrane. It has been shown that activation of AMPA receptors can evoke seizures in the preclinical models (
51). Based on previous studies, selective blockade of NMDA receptor alone is not sufficient to eliminate epileptiform discharges and AMPA receptors play a key role in modulating excitatory synaptic transmission in epilepsy (
52,
53).
In addition, we showed that inactivation of AMPARs in the somatosensory cortex suppressed seizure activities in the absence epileptic rats.
The anti-epileptic effect of AMPA receptor antagonists has been reported. These components noticeably reduce or suppress epileptiform activity by inhibiting glutamate-mediated excitation. It was shown in
in-vitro (
54-
57) as well as
in-
vivo animal models of seizure that AMPA receptor antagonists markedly reduce or abolish epileptiform activity (
17,
58).
Our findings clarify the potential therapeutic role of AMPARs for absence seizures and in the modulation of cortical neural hyperactivity.
Distribution of GluR1 in the somatosensory cortex of two- and six-month-old WAG/Rij and Wistar rats. (A) Photomicrographs of GluR1 distribution in the somatosensory cortex of two- and six-month-old WAG/Rij and Wistar rats. (B) The bar graph shows quantitative results of GluR1 distribution in the somatosensory cortex. GluR1 highly expressed in the both layers of somatosensory cortex in the 6-old-month WAG/Rij rats. ××× indicates p < 0.001 compared to Wistar rats. *** and ** indicate p < 0.01 and p < 0.001 respectively compared to 2-old-month WAG/Rij rats
The effect of AMPARs agonist and antagonist on the SWDs. (A) ECoG traces were recorded before, during and after drug administration. (B and C) Bar charts summarized mean ± SEM of SWDs amplitudes and durations. Micro injection of agonist significantly increased the duration (p < 0.001) and amplitude (p < 0.01) of SWDs in comparison with before injection. Administration of AMPARs antagonist on the somatosensory cortex reduced the amplitude (p < 0.001) and duration (p < 0.01) of SWDs in comparison with before injection. In 2 h after injection (recovery time) the amplitude and duration of SWDs in agonist (p < 0.05 for amplitude and p < 0.001 for duration) and antagonist (p < 0.01 for amplitude and p < 0.05 for duration) groups has changed significantly compared to injection time and reached near the initial values