DNA is a remarkable bioreceptor for a variety of small molecules and it can be considered as a major biological target to design anticancer agents. DNA-intercalating agents have been studied for several decades, and a few representatives (ellipticine, anthracyclines, acridines, and anthraquinones) are routinely used in the clinic for the treatment of cancers (
22). This study shows how different methods, such as UV-visible, fluorescence, viscosity, and competitive interaction between the NR dye and IBT with DNA, can be used to elucidate the binding of IBT to target DNA molecules.
Absorption spectra studies
Absorption spectra of IBT interaction with DNA
The electronic absorption spectroscopy is the most common way to investigate the interactions of compounds with DNA. A compound binding to DNA through intercalation usually results in hypochromism and bathochromism, due to the intercalation mode involving a strong π–π stacking interaction between an aromatic chromophore and the base pairs of DNA. It seems to be generally accepted that the extent of the hypochromism in the UV–visible band is consistent with the strength of intercalative interaction. The absorption spectra of IBT in the absence and presence of CT-DNA (at a constant concentration of IBT, [IBT] = 50 µM) are given in
Figure 2.
Absorption spectra of the IBT in 0.01 M Tris-HCl buffer (pH 7.4) at room temperature in the presence of increasing amounts of CT-DNA. [IBT] = 50 µM, [DNA] = 0–80 µM from top to the bottom. Arrows indicate the change in absorbance upon increasing the DNA concentration. Inset: plot of [DNA]/εa-εb vs. [DNA]
Titration of calf thymus DNA with the IBT caused some spectral changes. The bands at 356 nm showed considerable hypochromism, up to 22% and a small shift of the absorption maximum (from 356 to 357 nm) when saturated around [DNA]/[IBT] = 1.6.
Large hypochromism in the absorption bands of the IBT in the presence of double helical DNA is usually characteristic of intercalation of molecule into DNA base pairs, due to the strong stacking interaction between the aromatic chromophore and the base pairs (
23). So, the above phenomenon is indicative of most probable intercalative binding mode of IBT with calf thymus DNA (
24,
25). It should be noted that no significant effect on the absorption bands of the molecule in the presence of double helical DNA, is characteristic of a groove binder.
For further study of IBT interaction with DNA, the intrinsic binding constant between mentioned molecule and DNA was calculated; the intrinsic binding constant, Kb for IBT with CT-DNA was determined according to the following equation:
[DNA]/(εa-εf) = [DNA]/(εb-εf) + 1/Kb(εb-εf) Equation(1)
Where [DNA] is the concentration of DNA in base pairs. ε
a, ε
f and ε
b are the apparent, free and bound complex extinction coefficients, respectively. In particular, ε
f was determined by a calibration curve of the isolated IBT in aqueous solution, following Beer’s law. ε
a was determined as the ratio between the measured absorbance and the IBT concentration, A
obs/ [IBT]. A plot of [DNA]/(ε
a-ε
f) vs. [DNA] gives a slope of 1/( ε
b-ε
f) and a y-intercept equal to 1/K
b (ε
b-ε
f); K
b is the ratio of the slope to the y-intercept (Inset in
Figure 2). The K
b value was calculated to be 1.03×10
5 M
-1 (
24). In the previous study (
13), a K
b value of 7.32×10
4 M
-1 was determined (using UV) for isatin (a groove binder). The binding constants indicate that IBT binds more strongly than isatin. This result is expected, since IBT possesses greater planer area and extended π system than that of isatin, which will lead to IBT penetrating more deeply into, and stacking more strongly with base pairs of DNA (
26). These data indicate that the intercalated drug has a significant effect on the strength of DNA binding.
Absorption spectra of NR dye interaction with DNA
In general, if a small molecule interacts with DNA, changes in absorbance (hypochromism) and in the position of the band (red shift) should occur. Hypochromism and red shift are important evidences indicating that the small molecule has intercalated between DNA base pairs, and is involved in a strong interaction in the molecular stack between the aromatic chromophore and the base pairs (
27,
28). The spectral effects could be considered as follows: the empty π*-orbital of the small molecule couples with the π-orbital of the DNA base pairs, which causes an energy decrease, and a decrease of the π →π* transition energy. Therefore, the absorption of the small molecule should exhibit a red shift. At the same time, the empty π*-orbital is partially filled by electrons, reducing the transition probability, which leads to hypochromism.
In our work, UV–Vis spectra were recorded from solutions of the NR at constant concentration mixed with different concentrations of the DNA. In general, there is a maximum absorption at about 454 nm in the spectrum of an NR solution when DNA is present. It was found that the band absorbance at 454 nm decreased with an observed hypochromicity of 37%, and a small red shift (~11 nm) was evident with the increasing concentration of DNA. A band developed at approximately 500 nm with the increasing of DNA concentration, and has been designated to the NR-DNA complex. It increases in intensity with DNA, and has a bathochromic shift to 540 nm. An isosbestic point at 494 nm provided evidence of the new DNA–NR complex formation (
29,
33).
Absorption spectra of competitive interaction of IBT and NR with DNA
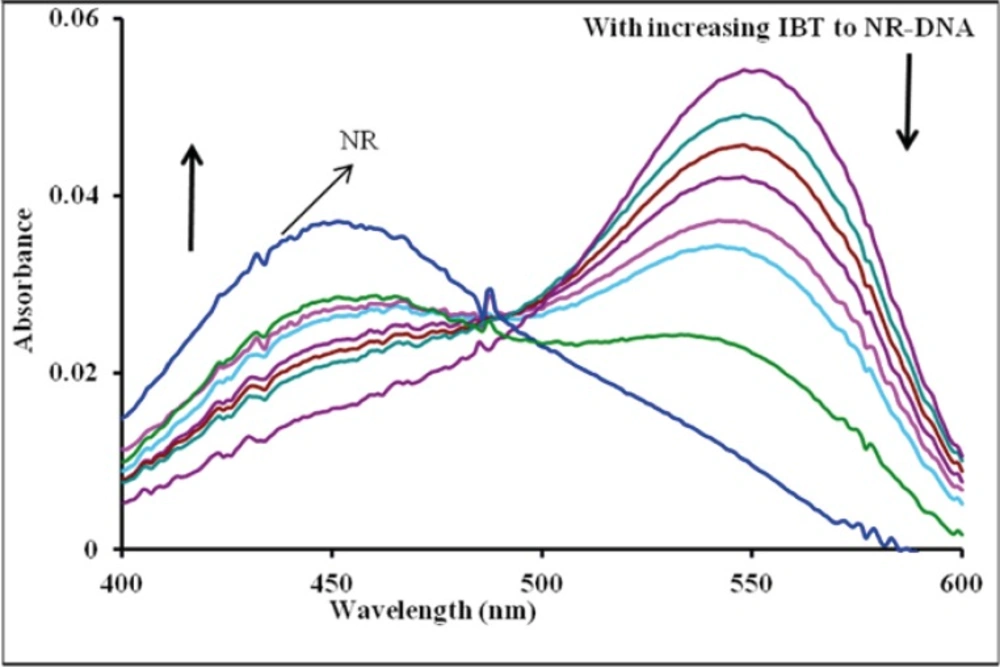
As shown in
Figure 3, with increasing the concentration of IBT, the maximum absorption at 540 nm of the NR-DNA spectrum decreases but a slight intensity increase is observed in the developing band at 454 nm. Comparing this observation to the behavior of the free NR absorption band at 454 nm in the presence of increasing concentration of DNA, it is noted that its intensity increases. The observed changes in intensity and position of the bands with increasing amounts of IBT added to the NR-DNA complex solution suggest that some of the NR molecules, which were intercalated into the DNA base pairs, were exchanged by IBT. This is consistent with the view that planarity of a molecule is one of the necessary conditions for efficient intercalation into the double helix (
30). An aromatic ring stacking between nucleobase pairs is regarded as a major driving force for binding of an intercalator into the double helix. Since the IBT molecule contains a planar aromatic ring (
Figure 1), which can stack between DNA bases, IBT should be able to intercalate into the double helix. Thus, considering the above results, it is evident that with the use of NR as an indicator, and UV–Vis spectrophotometry to probe the interaction between IBT and DNA under neutral pH conditions, the drug, IBT, binds to DNA by intercalating between the base pairs (
17,
29,
31).
Effect of IBT on absorption spectra of NR-DNA: CDNA = 8×10−5 M, CNR = 2.00×10−5 M, and CIBT = 0.0, 0.39, 1.5, 2.3, 3.4, 4.9, 7.5 and 11.5×10−5 M, corresponding to the curves, respectively
Fluorescence spectroscopic studies
Fluorescence spectra of IBT interaction with DNA
The IBT can emit luminescence in Tris buffer with a maximum wavelength of about 541 nm.
Figure 4 shows the emission spectrum of IBT in the absence and presence of varying amounts of DNA. As seen from the Figure, the intensity of emission at 541 nm increases appreciably in the presence of DNA. The enhancements of emission intensity imply that IBT molecules can insert between DNA base pairs. As a result, IBT molecules are protected from solvent water molecules by the hydrophobic environment inside the DNA helix; the accessibility of solvent water molecules to these compounds is reduced. The binding of IBT molecules to DNA leading to a marked increase in emission intensity also agrees with those observed for other intercalators (
34,
35).
Emission enhancement spectra of IBT (37 µM) in the absence (bottom spectrum) and presence of increasing amounts of calf thymus DNA (7.4, 18.5, 29.6, 37, 55.5, 66.6 and 74 µM; subsequent spectra). Arrow shows the emission intensity changes upon increasing DNA concentration
Characteristics of fluorescence spectra of interaction between IBT and DNA–NR
The interaction procedure of NR with CT-DNA at pH 7.4 was characterized by the fluorescence spectra (
32,
33). They showed that initially, NR produced only weak fluorescence in the Tris–HCl buffer due to quenching by the solvent molecules. However, with the addition of increasing concentration of DNA, fluorescence intensity of NR was enhanced substantially because of its intercalation into DNA base pairs. Such spectral effects are similar to those observed in the interaction of ethidium bromide (EB) and DNA (
28). This molecule is well known as a typical intercalator, and it has a weak fluorescence, which in the presence of DNA is significantly enhanced. This is attributed to strong intercalation of EB between the adjacent DNA base pairs (
24). In the present study, with the addition of IBT to a solution DNA–NR, some NR molecules were released into solution after an exchange with the IBT, and this resulted in fluorescence quenching together (
Figure 5A), indicating that interactions between DNA–NR and IBT occurred and DNA–IBT complex may form.
Fluorescence quenching refers to any process that is a decrease of the fluorescence intensity from a fluorophore due to a variety of molecular interactions. These include excited-state reactions, molecular rearrangements, energy transfer, ground-state complex formation, and collisional quenching. Quenching can occur by different mechanisms, which usually classified as dynamic quenching and static quenching. In general, dynamic and static quenching can be distinguished by their different dependence on temperature (
36). Dynamic quenching depends upon diffusion. Since higher temperatures result in larger diffusion coefficients, the bimolecular quenching constants are expected to increase with increasing temperature. In contrast, increased temperature is likely to result in decreased stability of complexes, and thus lower values of the static quenching constants (
19,
36-
39)
.The data obtained was analyzed by the Stern–Volmer equation (Equation 2) (
40):
F0/F = 1 + Kq τ0 [Q] = 1 + Ksv [Q] Equation (2)
Where F0 and F are the fluorescence intensities of DNA–NR in the absence and presence of the quencher, respectively. Kq is the quenching rate constant of the biomolecule, KSV is the Stern–Volmer quenching constant which can be considered as a measure for efficiency of fluorescence quenching by IBT, τ0 is the average lifetime of the biomolecule without quencher, and [Q] is the concentration of quencher (IBT).
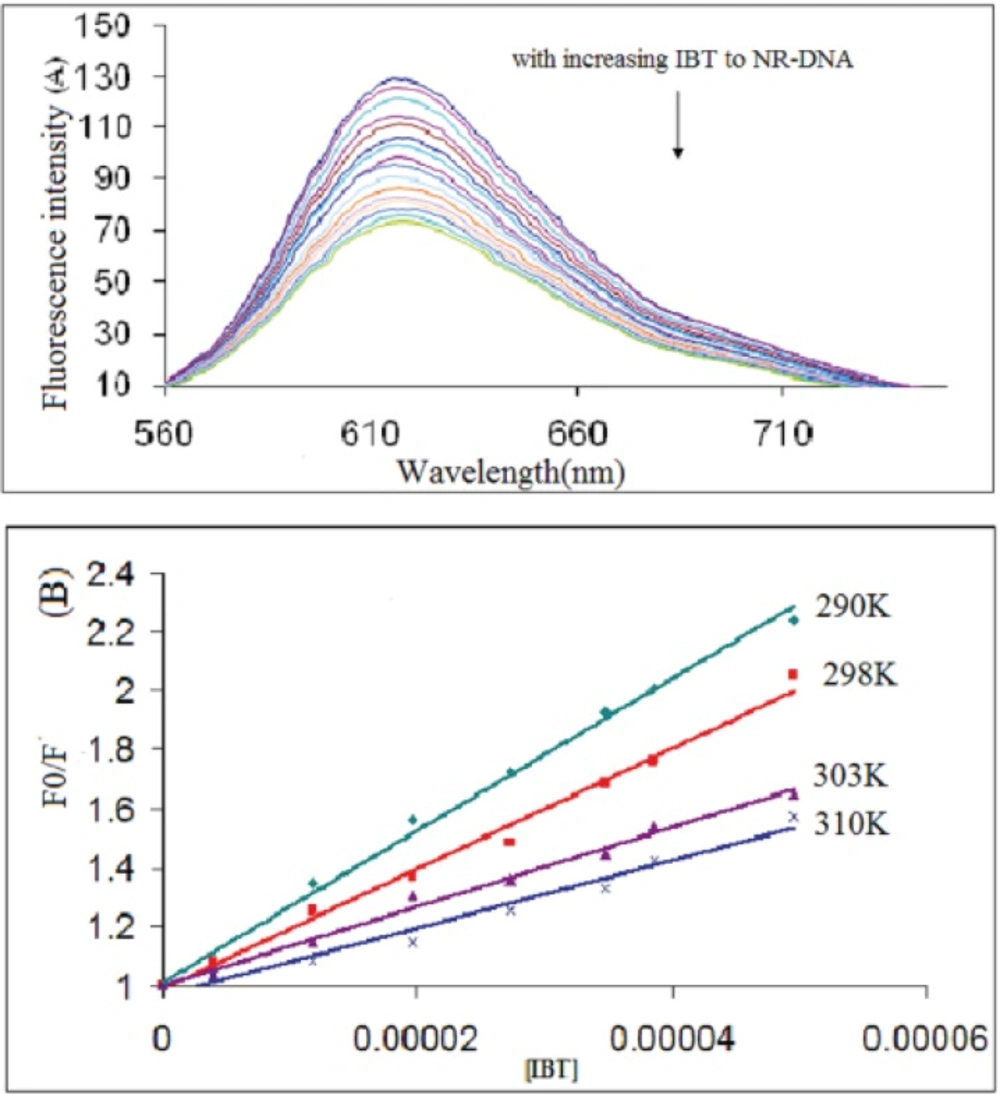
The K
sv of NR-DNA fluorescence by IBT at different temperatures (290, 298, 303, and 310 K) was obtained and the results are shown in
Figure 5B and indicated in
Table 1. These results show that IBT can quench NR-DNA fluorescence in a static quenching procedure, because the K
sv has been decreased by temperature rising (
33).
According to literature (
44) τ ≈10
-8 s, so the bimolecular quenching constant (K
q) was calculated to be 1.14×10
12 at 310 K. It is larger than the limiting diffusion rate constant of biomolecule (2.00×10
10), which indicates the static quenching occurred in the NR-DNA quenching by IBT (
37,
41).
(A) Fluorescence emission spectra of the competition between IBT and NR. CDNA = 9.0×10−5 M, CNR = 3.0×10−5 M and IBT = 0 - 4.9×10-5 M corresponding to the curves, respectively. (B) The Stern–Volmer plots for the quenching of DNA–NR by IBT at different temperatures.
| Temperature (K) | Ksv(L/mol)×104 | Kq(L/mol)×1012 | Kf | n | ΔG (kJ/mol) | ΔH(kJ/mol) | ΔS(J/molK) |
|---|
| 290 | 2.57 | 2.57 | 1.04×105 | 1.13 | -71.664 | -49.87 | -75.152 |
| 298 | 2.04 | 2.04 | 7.52×104 | 1.13 | -72.265 | | |
| 303 | 1.34 | 1.34 | 4.77×104 | 1.12 | -72.641 | | |
Binding constant and binding sites
As summed small molecules bind independently to a set of equivalent sites on a macromolecule, the equilibrium between free and bound molecules is given by the equation (
36,
37):
log(F0 − F)/F = log Kf + n log[Q] Eqution (3)
Where K and n are the binding constant and the number of binding sites, respectively. According to Equation (
3), the values of K
f and n can be obtained, which are summarized in
Table 1. The results show that the IBT binds to DNA with high affinity, which is much higher than that of isatin interacting with DNA, suggesting that the biological activities of IBT are superior to isatin (
13). The binding constant decreased with the rising temperature, which indicated that the binding is exothermic reaction.
The value of n approximately equal to 1 indicates the existence of just a single binding site in DNA for IBT, in other words the IBT molecules intercalate between all of the interbase spaces (
33,
42).
The thermodynamic parameters and nature of the binding forces
Considering the dependence of binding constant on temperature, a thermodynamic process was considered to be responsible for the formation of a complex. Therefore, the thermodynamic parameters dependent on temperatures were analyzed in order to further characterize the interaction forces between IBT and DNA. The interaction forces between a small molecule and macromolecule include hydrogen bonds, van der Waals force, hydrophobic force, electrostatic interactions,
etc. The thermodynamic parameters of binding reaction are the main evidence to confirm the binding force. If the enthalpy change (ΔH) does not vary significantly over the temperature range studied, then its value and that of entropy change (ΔS) can be determined from the van’t Hoff equation (
43):
log K = -ΔH/2.303RT + ΔS/2.303R Equation (4)
ΔG = ΔH - T ΔS Equation (5)
Where K is the binding constantat corresponding temperature and R is the gas constant. The temperatures used were 290, 298, 303 and 310K. The enthalpy change (ΔH) and entropy change (ΔS) were obtained from the slope and intercept of the linear van’t Hoff plot based on log K versus 1/T . The free energy change (ΔG) was estimated from Equation (5). The values of ΔH, ΔS and ΔG are listed in
Table 1. From
Table 1, it can be seen that the negative value of ΔG revealed the interaction process is spontaneous.
The free-energy changes (ΔG) for IBT–DNA interactions are negatively large due to their strong association. Ross and Subramanian reported that when ΔH<0 or ΔH≈0 and ΔS>0, the electrostatic force dominates the interaction; when ΔH<0 and ΔS<0, van der Waals interactions or hydrogen bonds dominate the reaction (
44); when ΔH>0 and ΔS>0, hydrophobic interactions dominate the binding process. Results indicated that ΔH<0 and ΔS<0. Therefore, van der Waals interactions or hydrogen bonds are the main forces in the binding of the investigated drug to CT-DNA, and the mode of binding is intercalation. In addition, the negative entropy change results from the intercalation of IBT between CT-DNA bases, accompanied by the loss of translational and rotational degrees of freedom (
45-
47). It also should be added that groove binding is predominantly entropically driven, whereas intercalation is enthalpically driven (
46).
Viscosity measurements
To further clarify the nature of the interaction between the IBT and DNA, viscosity measurements were carried out by varying the concentration of the added IBT to DNA solution. The specific viscosity of the DNA sample clearly increases with the addition of the IBT (
Figure 6). The viscosity studies provide a strong evidence for intercalation. The viscosity increase of DNA is attributed to the intercalative binding mode of the IBT, due to effective DNA length increase (
48,
49).
A classical intercalative mode causes a significant increase in viscosity of DNA due to an increase in separation of base Pairs at intercalation sites, and hence an increase in overall DNA length happens. By contrast, complexes that bind exclusively with the DNA grooves by partial and/or non classical intercalation, under the same conditions, typically cause less positive or negative or no changes in the DNA solution viscosity, for example isatin causes no significant changes in the DNA viscosity (
35,
50,
51).
Effect of increasing amounts of IBT on the viscosity of calf thymus DNA (5× 10 -5M) in 10 mM Tris–HCl buffer (pH 7.4) at 298K (ri = [IBT]/[DNA] = 0.0, 0.5, 1, 1.2, 1.5, 1.8, and 2).
Circular dichroism spectral studies
Circular dichroism (CD) spectroscopy is a very practical method to analyze the structure of optically active materials, such as DNA and proteins; therefore, the CD technique was used to determine the DNA conformational changes. In addition CD signals are quite sensitive to the mode of DNA interaction with small molecules.
The CT-DNA in the B conformation shows two conservative CD bands in the UV region; a positive band at around 275 nm (due to base stacking) and a negative band at around 245 nm (due to polynucleotide helicity) (
52). In
Figure 7, the changes in the CD spectrum of CT-DNA in presence of increasing concentrations of IBT are depicted. The positive band showed increase in molar ellipticity without any significant shift in the band maxima when the IBT concentration was progressively increased. This observation is supportive for the intercalative mode of binding of the IBT, where the IBT molecules stack in between the base pairs of DNA thus leading to the enhancement in the positive band (
53). It is also evident from the CD spectrum that the binding of the IBT leads to decrement in helicity and consequently change in the conformation of CT-DNA, shifting from a more B-like to a more A-like DNA, which is indicative of intercalative binding mode of mentioned molecule to DNA molecule (
44).
Circular dichroism spectra of DNA (8.0×10-5 M) in 10 mM Tris-HCl buffer, in the presence of increasing amounts of IBT (ri = [IBT]/[DNA] = 0.0, 0.05, 0.4, and 0.7).

![Absorption spectra of the IBT in 0.01 M Tris-HCl buffer (pH 7.4) at room temperature in the presence of increasing amounts of CT-DNA. [IBT] = 50 µM, [DNA] = 0–80 µM from top to the bottom. Arrows indicate the change in absorbance upon increasing the DNA concentration. Inset: plot of [DNA]/εa-εb vs. [DNA]](https://brieflands.com/journals/ijpr/articles/125264/figures/ijpr-14-111-g002-preview.webp)
![Effect of increasing amounts of IBT on the viscosity of calf thymus DNA (5× 10<sup> -5</sup>M) in 10 mM Tris–HCl buffer (pH 7.4) at 298K (ri = [IBT]/[DNA] = 0.0, 0.5, 1, 1.2, 1.5, 1.8, and 2).](https://brieflands.com/journals/ijpr/articles/125264/figures/ijpr-14-111-g006-preview.webp)
![Circular dichroism spectra of DNA (8.0×10<sup>-5</sup> M) in 10 mM Tris-HCl buffer, in the presence of increasing amounts of IBT (ri = [IBT]/[DNA] = 0.0, 0.05, 0.4, and 0.7).](https://brieflands.com/journals/ijpr/articles/125264/figures/ijpr-14-111-g007-preview.webp)
