
Introduction
Experimental
Results
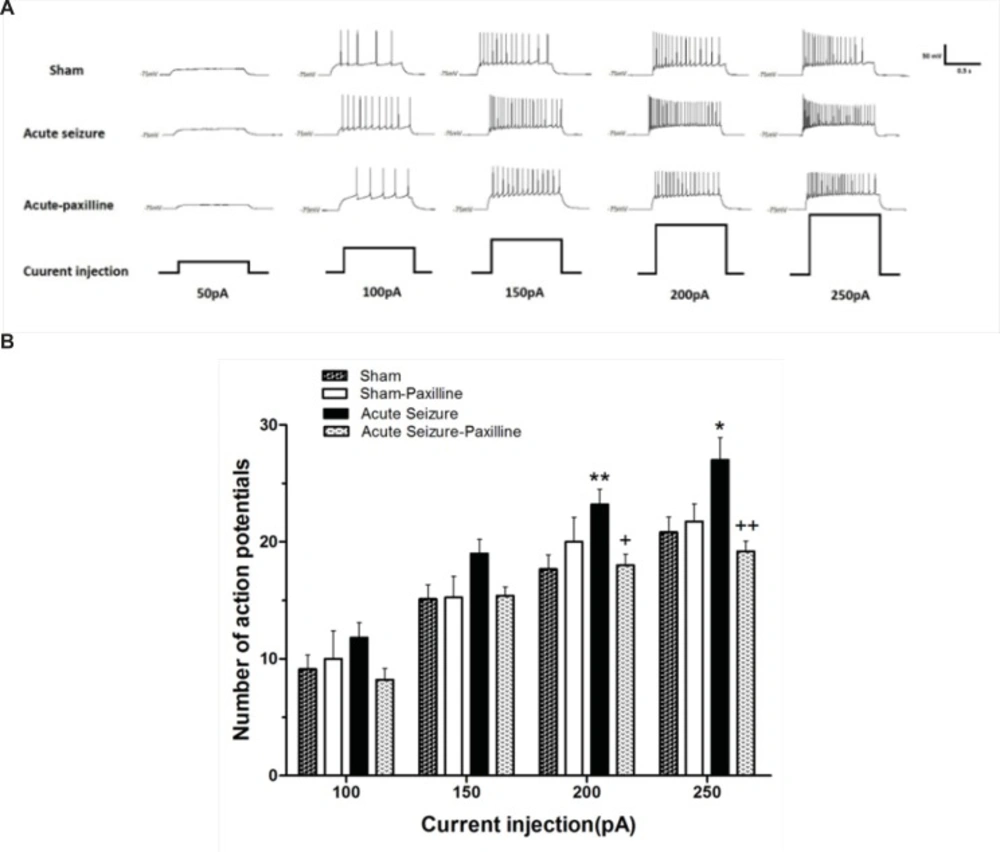
Alteration in the firing rate of dentate gyrus GCs recorded 24 h after pilocarpine-induced seizure. (A) Representative traces show the differences in firing rate of the GCs in response to 1000 ms depolarizing pulses from 50 pA to 250 pA in sham (above), acute seizure (middle) and acute seizure + paxilline (down) groups. (B) Depolarizing current injection ranging from 50 pA to 250 pA increased the number of APs in seizure group compared to sham group. The application of paxilline (1 μM) reversed pilocarpine-induced increase in APs. Data were shown as mean+SEM (N = 10).
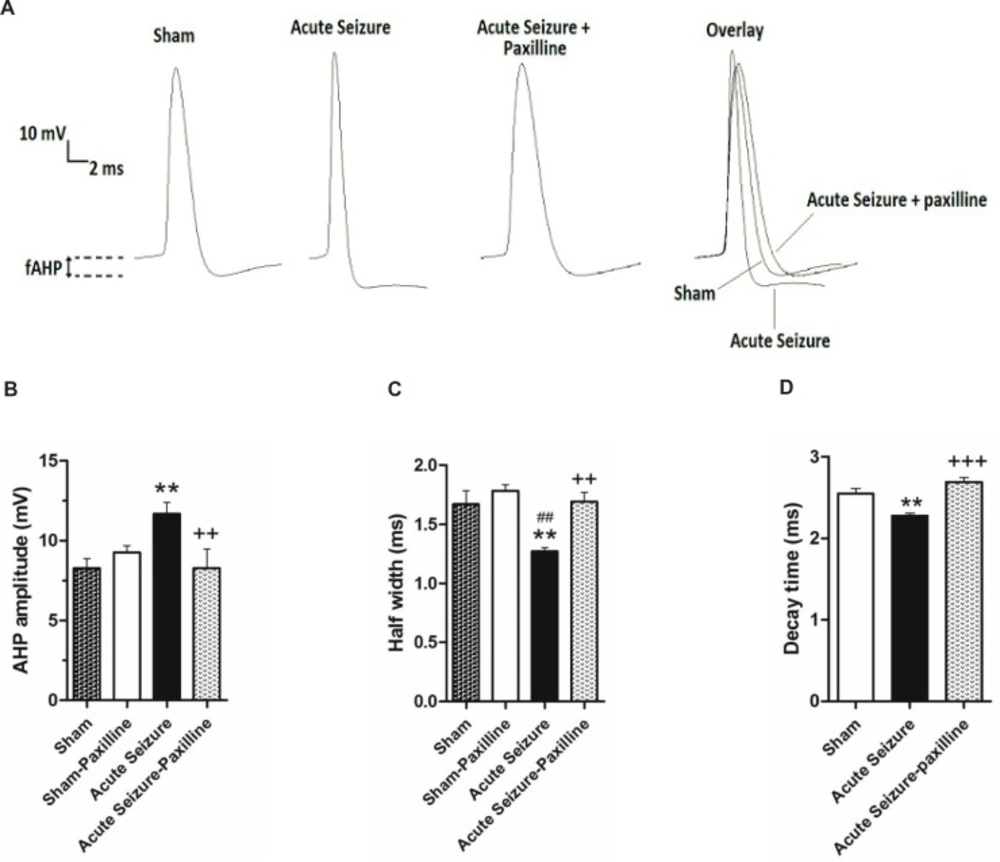
Pilocarpine-induced changes in AHP amplitude and AP half-width were reversed by bath application of paxilline. (A) Representative trace of 1st AP of sham, acute seizure and acute seizure + paxilline groups during a train of AP evoked by 200 pA current injection for 1000 ms. (B) fAHP amplitude was measured from the level of RMP to the peak of the hyperpolarization. fAHP amplitude significantly increased during acute phase of pilocarpine-induced seizure compared to sham group. Paxilline decreased fAHP amplitude to the sham values. (C) AP half- width significantly decreased during acute phase of seizure. Paxilline increased AP half-width to normal values and reversed the effect of seizure on AP half-width. (D) The decay time of AP was increased in pilocarpine-induced seizure group and was reversed after bath application of paxilline.
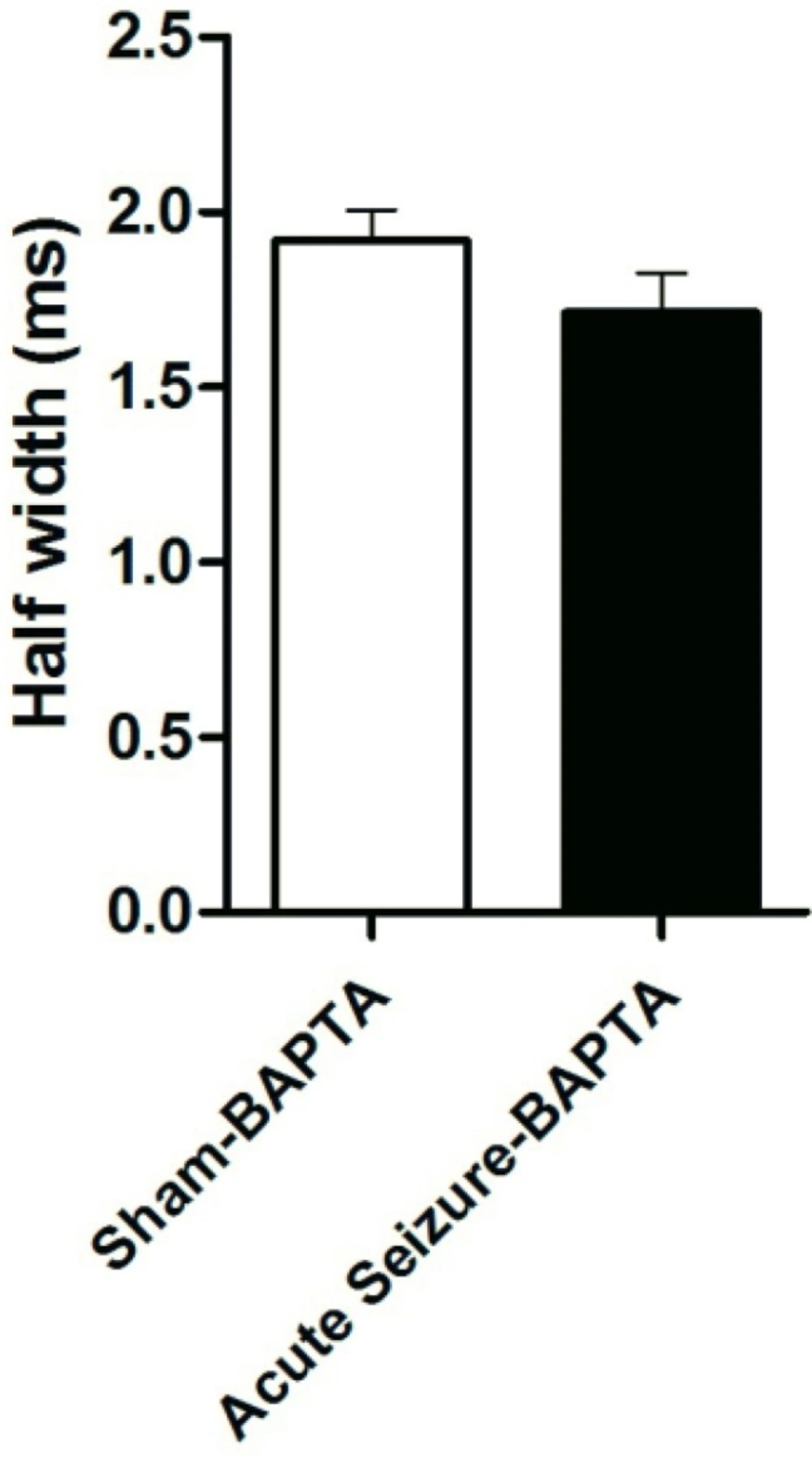
Effect of BAPTA on AP half-width. Application of BAPTA prevented change in AP half-width in seizure group compared to that of BAPTA-sham cells. Data were shown as mean + SEM (N = 5).
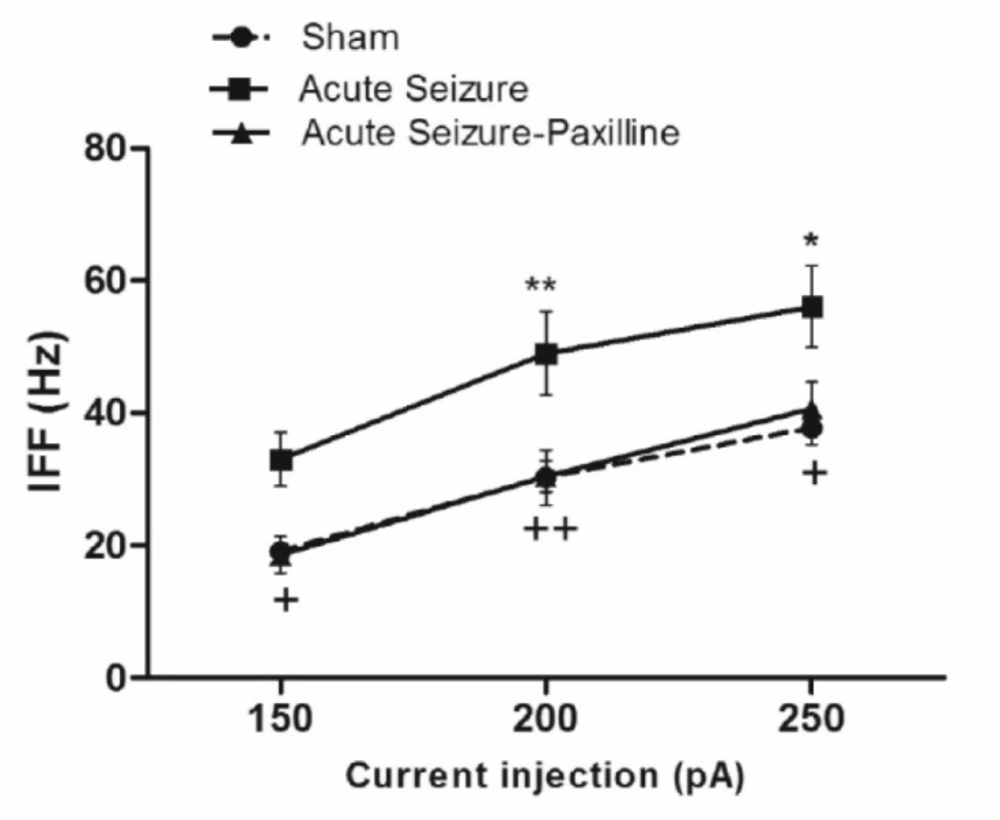
Seizure-induced increase in instantaneous firing frequency (IFF) was reversed by paxilline application. IFF was measured as the reciprocal of interval between the first and the second AP during 200 pA current injection. IFF significantly increased during acute phase of pilocarpine-induced seizure. After bath application of paxilline, IFF returned to sham levels. Data were shown as mean ± SEM (N = 6 in each group).
