Ten
gyrA mutants and their parent strain MG1655 were assessed for organic solvent tolerance. Results are shown in
Table 2. As can be seen from this table, MG1655 can grow in presence of hexane, but not cyclohexane. This is consistent with the previous result for this strain (
15). All mutants derived from this strain also show nearly the same results. Some mutants had slightly better growth on hexane.
To assess whether
gyrA mutants may acquire resistance to other antibiotics, the MIC of Tc for control strain and
gyrA mutants were measured. For MG1655, MIC was 3 µg/mL (
Table 2). This is consistent with previous data (
19). MICs of
gyrA mutants were nearly the same as that of MG1655, except some seemed to show better growth at 3 µg/mL tetracycline and their MICs were 4 µg/mL (
Table 2). Collectively, these results suggest that
gyrA mutants may either not acquire a mutation in
marR gene, or some of them acquire a mutation, but cannot cause high level of organic solvent tolerance and partial resistance to other antibiotics at once and need to be induced in the presence of inducers, such as tetracycline (
19).
| Strain/mutant | MIC* (μg/mL) |
|---|
| MG1655 wild type | 3 |
| W10-W11 | 3 |
| W25-W26 | 4 |
| W44-W45 | 3 |
| W47-W48 | 3 |
| W49 | 4 |
| W50 | 3 |
To verify this
marR gene and its upstream region,
marO in MG1655 and its 10
gyrA mutants were amplified.
Figure 2 shows the result of gel electrophoresis of the
marOR PCR product of MG1655. The same results were obtained for all
gyrA mutants. Then, the PCR products were sequenced using forward and reverse primer. Finally, the sequences were compared with the published
marR sequence of MG1655 strain, using EBI bioinformatics program for nucleotide pair wise alignment. MG1655 showed the complete match with previous published sequence for
marR (data not shown). Moreover, 7
gyrA mutants also showed the same result as MG1655. Their Marbox and promoter site were also intact. This suggests that their higher ciprofloxacin MICs may not be associated to the induction of drug efflux. However, 2 mutants with higher Tetracycline MIC, had mutations in
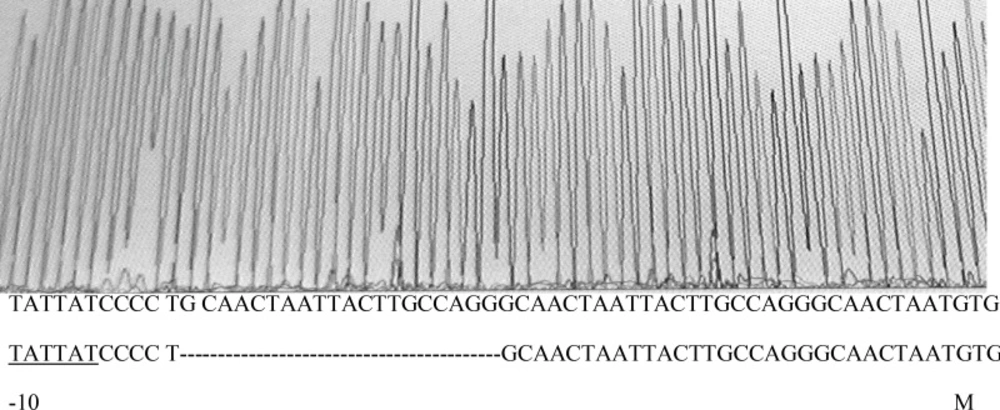
marOR. One contained a T→C change at nucleotide position 221 in coding region of
marR that alters methionine-74 to threonine (
Figure 3), but its marbox and promoter site was without change. The other mutant harbors, a 20-base pair tandem duplication of a sequence (GCAACTAATTACTTGCCAGG) started 6-base pair downstream from the 10-position of promoter site (
Figure 4). This type of mutation was described before (
14). However, its marbox was intact. The last mutant with higher tetracycline MIC did not possess a mutation in
marOR. This suggests that the reason of slightly different tetracycline MIC may be the occurrence of mutation in a gene that encodes a suppressor for the pump.
In the previous study, ciprofloxacin-resistant mutants which possess mutations in
gyrA were isolated (
7). These
gyrA mutants had different levels of resistance to ciprofloxacin. It was explained that one reason for this difference is due to the decreased levels of drug accumulation (
6). The intracellular amount of antibiotic can be regulated through a efflux pump, AcrAB-TolC (
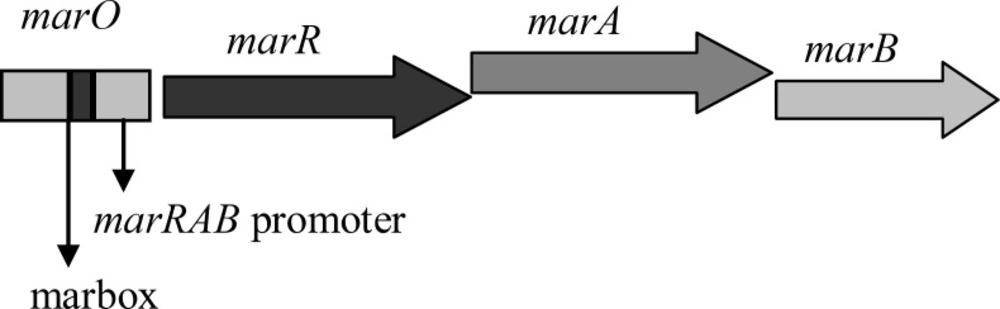
10). The activation of this pump is associated with the presence of MarA, the transcription activator protein. The gene that encoded this protein is located in
marRAB operon, consisting
marR,
marA and
marB genes. This operon is normally inactivated through the product of
marR gene, MarR. The inactivation of MarR is caused through the mutations in
marR (
17). This leads to the expression of a multidrug resistance phenotype (MDR). This phenotype is also associated to the high level of organic solvent tolerance (
15,
16). Therefore, in this study, ciprofloxacin-resistant mutants with different MICs were examined for the tolerance of organic solvents, resistance to tetracycline and presence of
marR mutation.
Gel analysis of PCR product. Lane M and A contain 1 Kb DNA ladder and PCR product, respectively.
The obtained results revealed that none of the gyrA mutants even those with higher ciprofloxacin MICs possess the MDR phenotype de novo, but some gyrA mutants may gain this phenotype gradually following the exposure to an inducer of marRAB operon, such as tetracycline. This was revealed through the discovery that 3 out of 10 gyrA mutants are slightly more tolerant to hexane and resistant to tetracycline compared to MG1655.
Sequence output from PCR product of gyrA mutant using forward and reverse primers. On the bottom of graph, the first and second rows of nucleotide sequences belong to mutant and wild type strain, respectively. The third line shows the amino acid sequence of that region in wild type strain.
Our finding that 7
gyrA mutants are the same as MG1655 for sensitivity to tetracycline, tolerance of organic solvent and devoid of
marOR mutation, implies that the reason for their higher ciprofloxacin MICs in comparison with MG1655 may be the presence of mutations in
gyrB,
parC or
parE (
6). The last two genes encode the subunits of topoisomerase IV, the minor target of fluoroquinolone antibiotics in
E. coli (
6).
Moreover, an
E. coli mutant (OST3408) was found by Asako that is tolerant not only to hexane, but also to cycloheexane (
15). Most of
E. coli strains are sensitive to cyclohexane. This mutant contains a substitution of serine for arginine at position 73 in the coding region of marR. The MarR protein has been shown to contain the helix-turn-helix motif started from the 60
th amino acid codon and extended to the 80
th one (
20). This motif is necessary for the binding of repressor to the promoter site. However, we found that a T→C mutation in marR that causes a substitution of threonine for metionine at position 74 does not confer tolerance to cyclohexane. This implies that both the position of amino acid and the type of substitutions in amino acid sequence of this motif are important on functionality of repressor. On the other hand, the alteration of the repressor binding sites interferes with repressor activity. We found a
marR mutation with altered repressor binding sites that has already been discovered (
14). It was found that this kind of mutation increases the activity of
marRAB operon (
14).
Sequence output from PCR product of gyrA mutant using forward and reverse primers. On the bottom of graph, the first and second nucleotide sequences belong to mutant and wild type strain, respectively. The -10 region of promoter, duplicated sequence and start codon are shown
Moreover, mutations in
acrR gene, encoding the repressor of AcrAB-TolC pump called AcrR, increase the activity of the pump (
21). This fact that one of the three slightly resistant mutants did not possess a mutation in
marOR, implies that this
gyrA mutant may acquire a mutation in
acrR gene.
Furthermore, highly organic solvent tolerant mutants gain mutations both in
marOR and
acrR (
22). Therefore, it is possible that our
gyrA marR double mutants are not resistant to cyclohexane due to their lack of a mutation in
acrR.