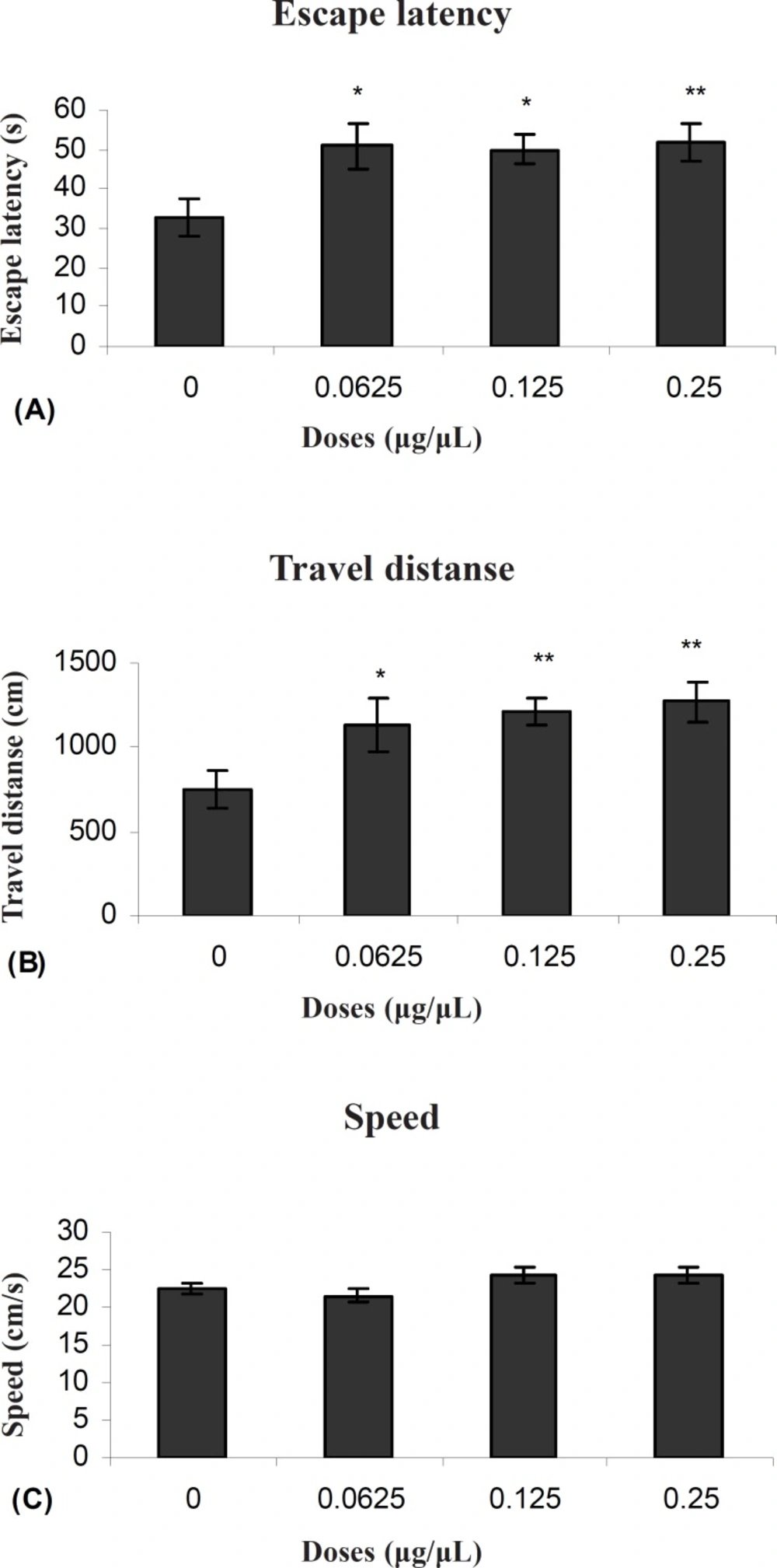
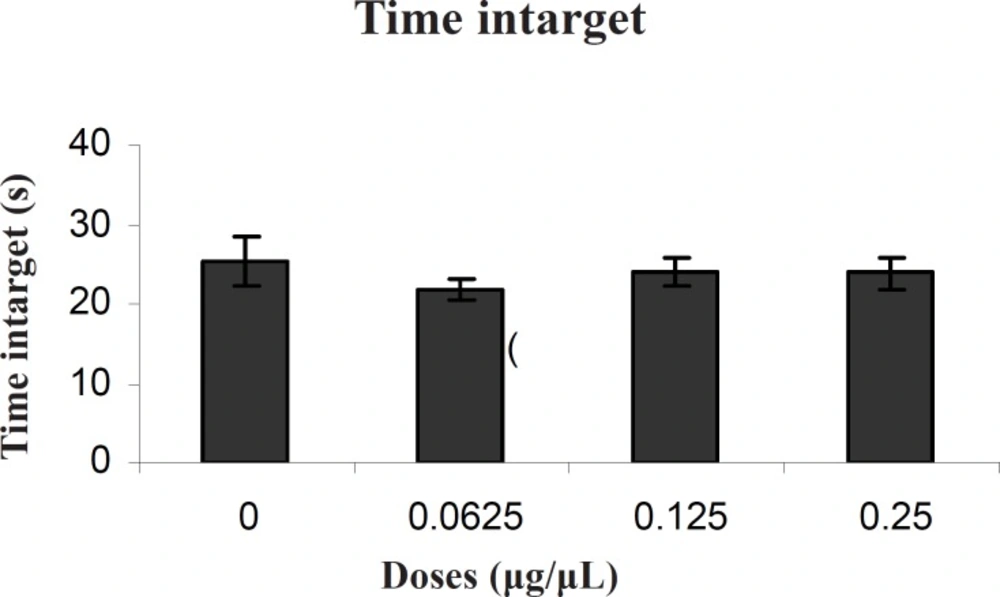
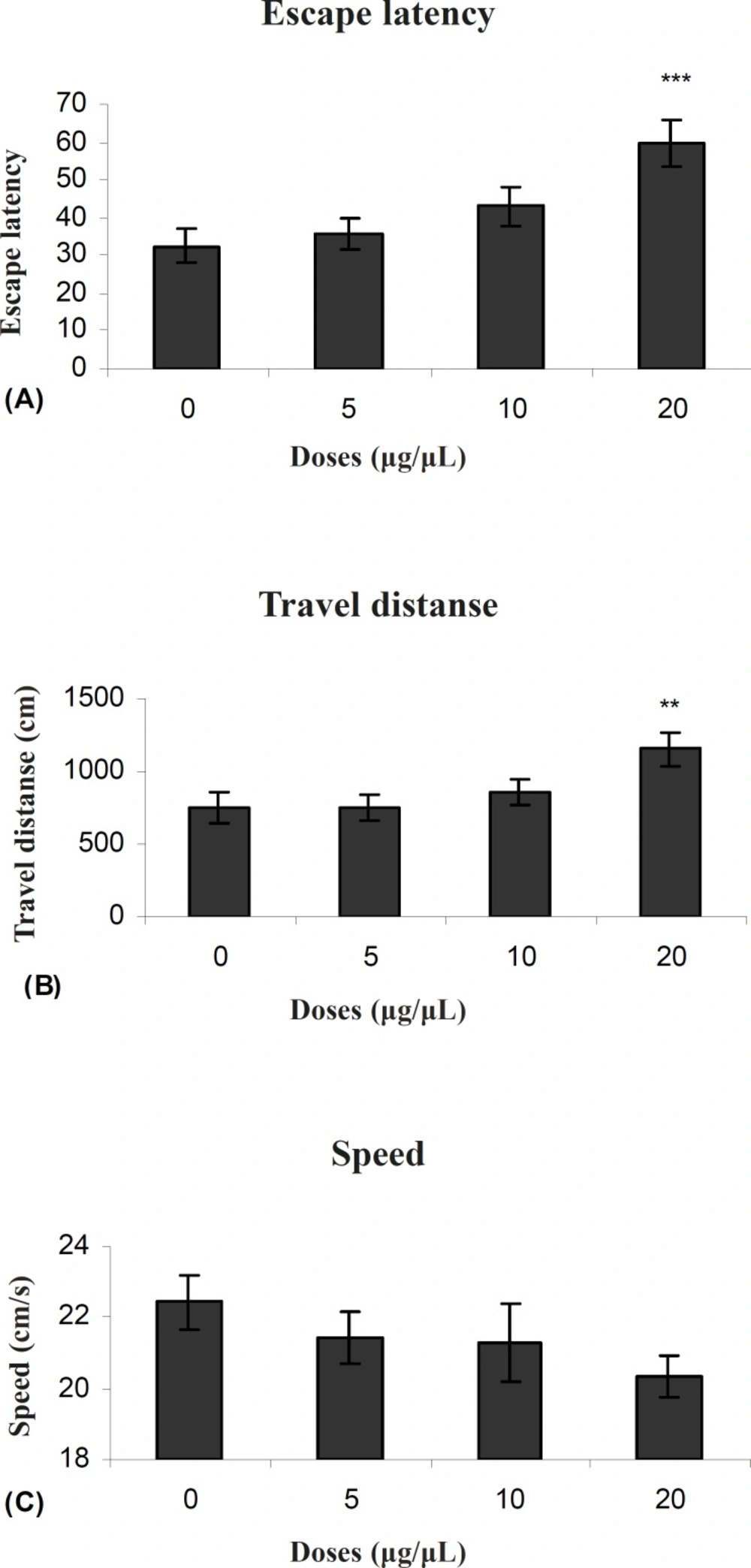
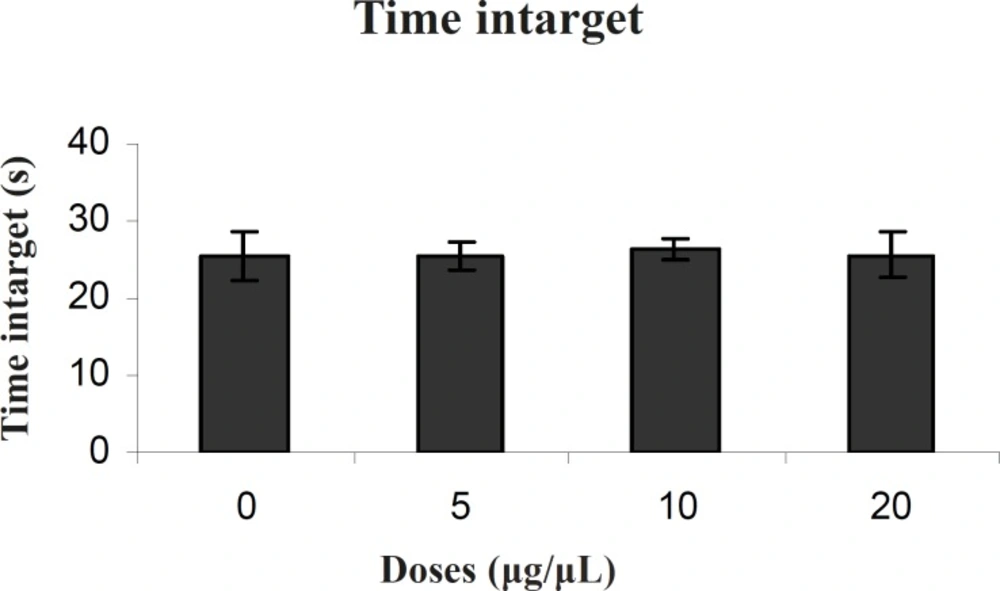
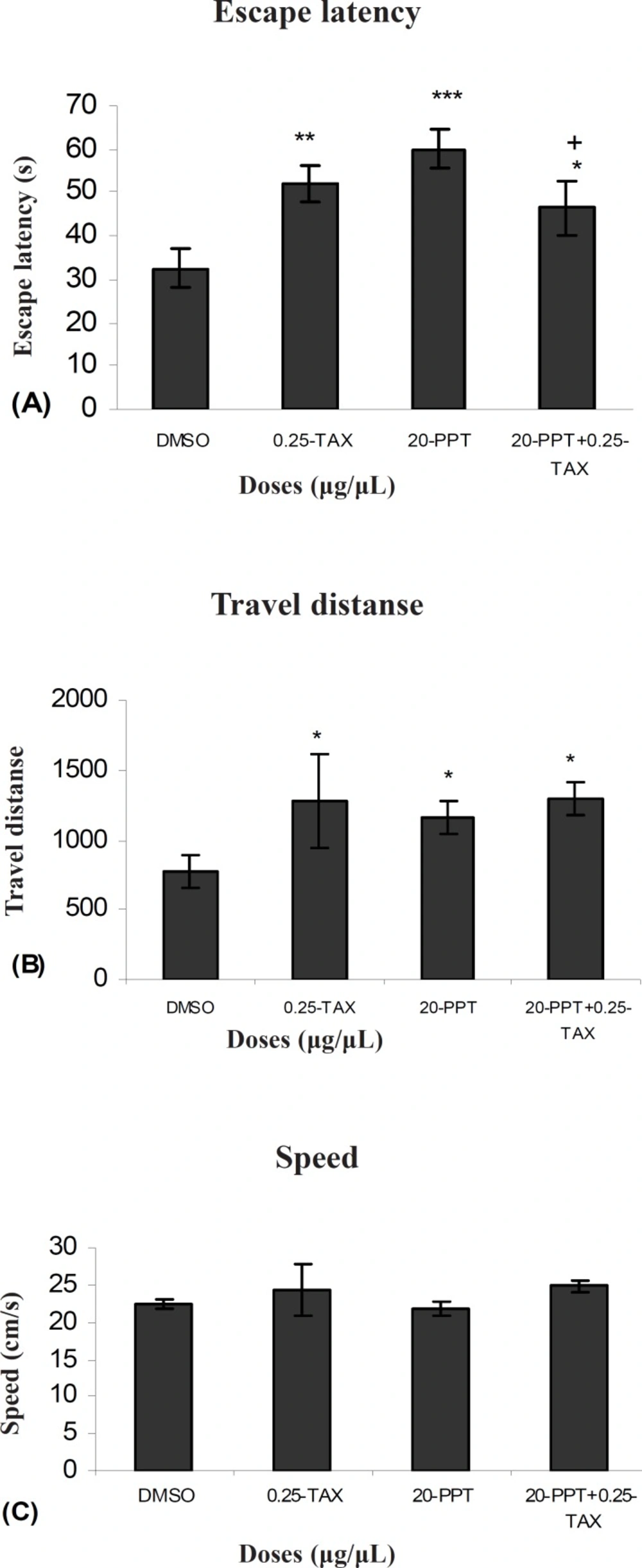
Our results showed that TAX (0.25 μg/0.5μl) as an ERs antagonist and PPT (20μg/0.5μl) as an ERα selective agonist significantly increased escape latency and travel distance in comparison to control group.
These results showed that PPT and TAX could impair acquisition of memory in water maze task. Also the results showed that TAX (0.25μg/0.5μl) could eliminate PPT (20μg/0.5μl) - induced impairment. In all experiments, there were no significant differences in swim speed indicating that there were no significant differences in sensory or motivational processes between the groups.
The mechanism of action of TAX on impairing learning and memory functions has not been clearly evaluated (
26). Estrogen replacement induces a 30% rise in both NMDA receptors and spines in the hippocampus of ovariectomized female rats (
26). Thus, one may speculate that TAX could affect the function of hippocampus to impair the memory ability (
27). Because TAX is an estrogen receptor antagonist in the central nervous system (
26), it could be assumed that this activity is the main cause of their memory impairing action.
A close interaction between estrogen and cholinergic function in the central nervous system has been reported (
28-
30). It has been inferred that estrogen affects learning and memory behaviors by modulation of basal cholinergic function (
27). For example, it is reported that steroidal sexual hormones can affect on acetylcholine transferase and acetylcholine esterase activities. The long-term memory is facilitated with estradiol in the one-trial passive avoidance conditioning test in adult male Wistar rats (
31). In ovariectomized rats, high-affinity choline uptake is reduced in the hippocampus and in the frontal cortex by 24% and 34%, respectively. This decline in high-affinity choline uptake is associated with a significant decrease in total avoidance in the active avoidance tests. Estrogen administration could reverse the effects of ovariectomy on high-affinity choline uptake in active avoidance and spatial memory behavior (
32). These data suggest that cholinergic neurons are estrogen-responsive and that continuous exposure to ovarian steroid is needed to maintain the normal memory function. Since TAX is an antiestrogenic drug, it is reasonable to assume that TAX - induced impairment memory might be due to the blockade of the estrogen actions, which subsequently affects the activity of the cholinergic system (
28).
Other mechanisms of action of TAX on memory function could not be excluded. It has been shown that TAX has a wide variety of pharmacological activities, such as the inhibition of protein kinase C (
33), acting as a calmodulin antagonist (
34), and acting as a histamine antagonist (
35). These TAX actions may directly or indirectly affect on memory function (
27).
Our results also showed that PPT (20 μg/0.5 μl) as an ERα selective agonist impaired acquisition of memory in Morris Water Maze task. Thus, it is possible that estrogen impairs memory via ERα. There are two possibilities for this finding:
1- Regulation of spinogenesis is one of the important estrogen roles in memory processes via producing new spines for creating new neuronal contacts (
36). Dendritic spine morphology analysis demonstrated that the density of thin type spines was selectively increased in CA1 pyramidal neurons within 2 h after application of 1 nM estradiol (
21).
Sometimes the spine density is decreased by the estradiol treatment. The estradiol-induced spinogenesis is region specific and heterogeneous. In fact, in CA3 pyramidal neurons, the total density of thorns of thorny excrescences (spine-like postsynaptic structures in the stratum lucidum of CA3, having contacts with mossy fiber terminals originated from granule cells) decreased dramatically to approximately 70% upon a 2-h treatment of 1 nM estradiol (
37). PPT significantly decreased the density of thorns from 2.19 to1.66 thorns/μm. Therefore, it may be PPT-induce impair memory processes by decreasing of spine density.
2- In memory processing, LTP, long term potentiation, (memory forming mechanism) and LTD (long term depreciation) are essential. Mutant mice, which show enhanced LTP and suppressed LTD, have shown impaired learning of Morris water maze (
38). This suggests that LTD may be required to “correct” wrong memories formed by initial LTP processes, which store not only correct information but also wrong information. (
21,
39). Investigations using specific estrogen agonists indicated that the contribution of ERα (but not ERβ) was essential to these estradiol effects. PPT at 100 nM exhibited a significant LTD enhancement in CA1 (
39). Therefore, it may be suggested that PPT impaired learning and memory via LTD enhancement and derangements in “correct” wrong memories formed by initial LTP processes.
In addition, our study showed that TAX as ERs antagonist (0.25 μg/0.5μl) could impair acquisition in MWM task. Also, our results showed that using TAX (ERs antagonist, 0.25 μg/0.5 μl) with ERα selective agonist (PPT (20 μg/0.5 μl) ) could eliminate impairment of using of PPT. It is demonstrated that estrogen modulates memory function via estrogenic receptors, especially ERα. Because PPT (agonist of ERα) in present of TAX (antagonist ERs) also shows an impairment effect on spatial learning and memory. Thus, it is possible that TAX has higher affinity to bind ERα than ERβ. Fugger N. et al. (2000) showed that ovariectomized (OVX) estrogen receptor-α-knockout (ERαKO) mice displayed impaired performance on the inhibitory avoidance (IV) task. In contrast to ERαKO mice, IA performance by OVX estrogen receptor-β-knockout (ERβKO) mice was not compromised. The results indicate an important role for ERα, relative to ERβ, in the establishment of cognitive function (
20). Only the estrogen receptor (ER) α-agonist, propyl pyrazole trinyl tris-phenol (PPT), induced the same enhancing effect as estradiol on spinogenesis in the CA1. The ERbeta agonist, (4-hydroxyphenyl)-propionitrile (DPN), did not affect on spinogenesis (
21). Regulation of spinogenesis is one of the important estrogen roles in memory processes via producing new spines for creating new neuronal contacts (
36). It demonstrates that probably ERα is more important than ERβ in learning and memory process. Other studies have shown that ERβ is an important modulator for learning and memory (
9,
24). Considering downregulation of ERα in the hippocampus and amygdalohippocampal area in estrogen-treated knockout individuals, it was proposed that ERβ influenced spatial memory by binding estradiol and preventing this downregulation, a mechanism that had previously been postulated for other areas of the brain (
9). Therefore, one could say that there are many of unknown proceeding in basis of learning and memory, also involvement of estrogenic receptors types in this process.
In summery, it seems that intra hippocampal injections of TAX as ERs antagonist and PPT as ERα selective agonist could impair spatial learning and memory in MWM task. Also results showed that TAX could eliminate impairment of memory by injection of PPT. This finding suggests that estrogens act in learning and memory via ERs, specifically ERα. In this process many different mechanisms involved such as, distinct differences in binding and transactivation properties of α- and β-receptors, interaction between estrogen and cholinergic function in CNS, spinogenesis, LTP (long term potentiation), LTD (long term depreciation), ERs particularly ERα .