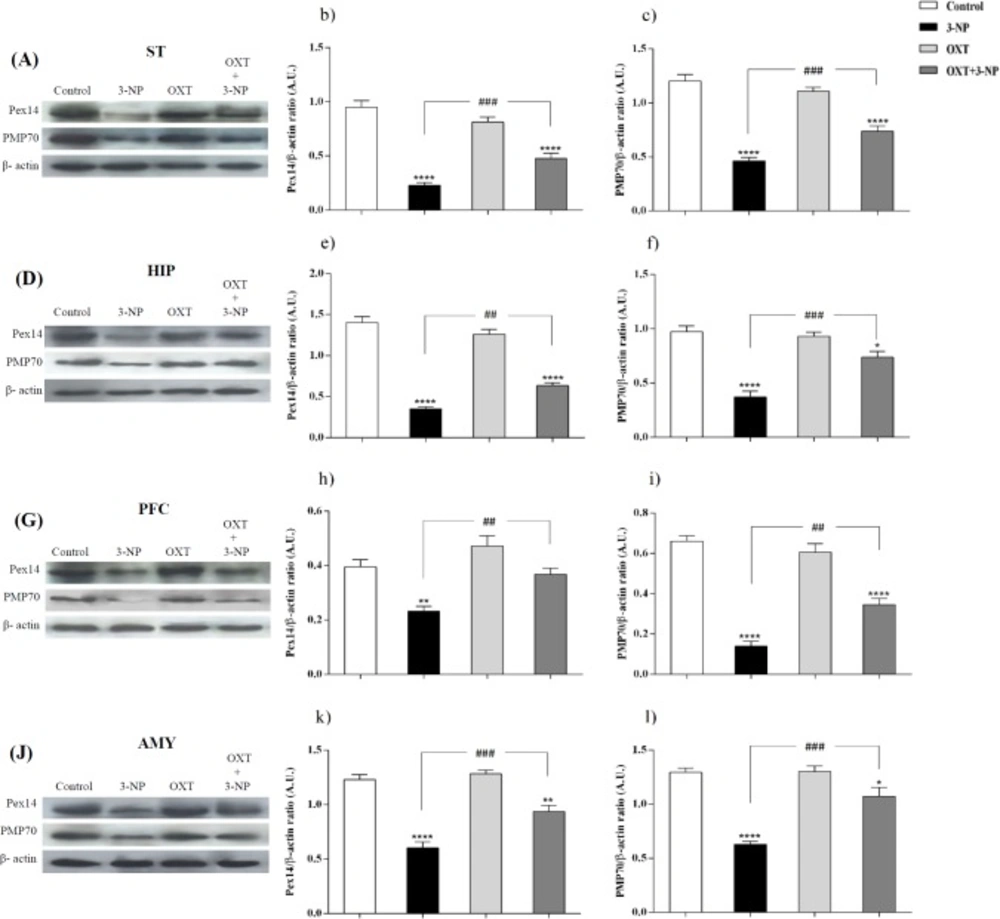
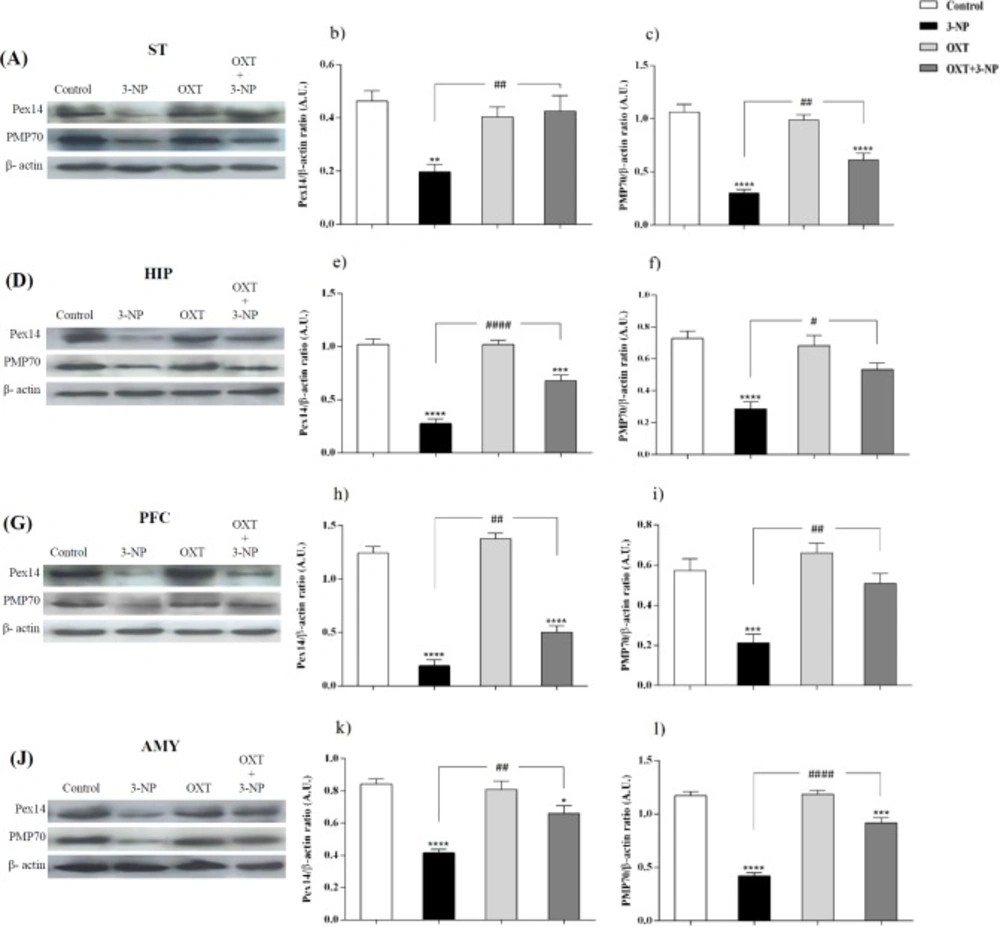
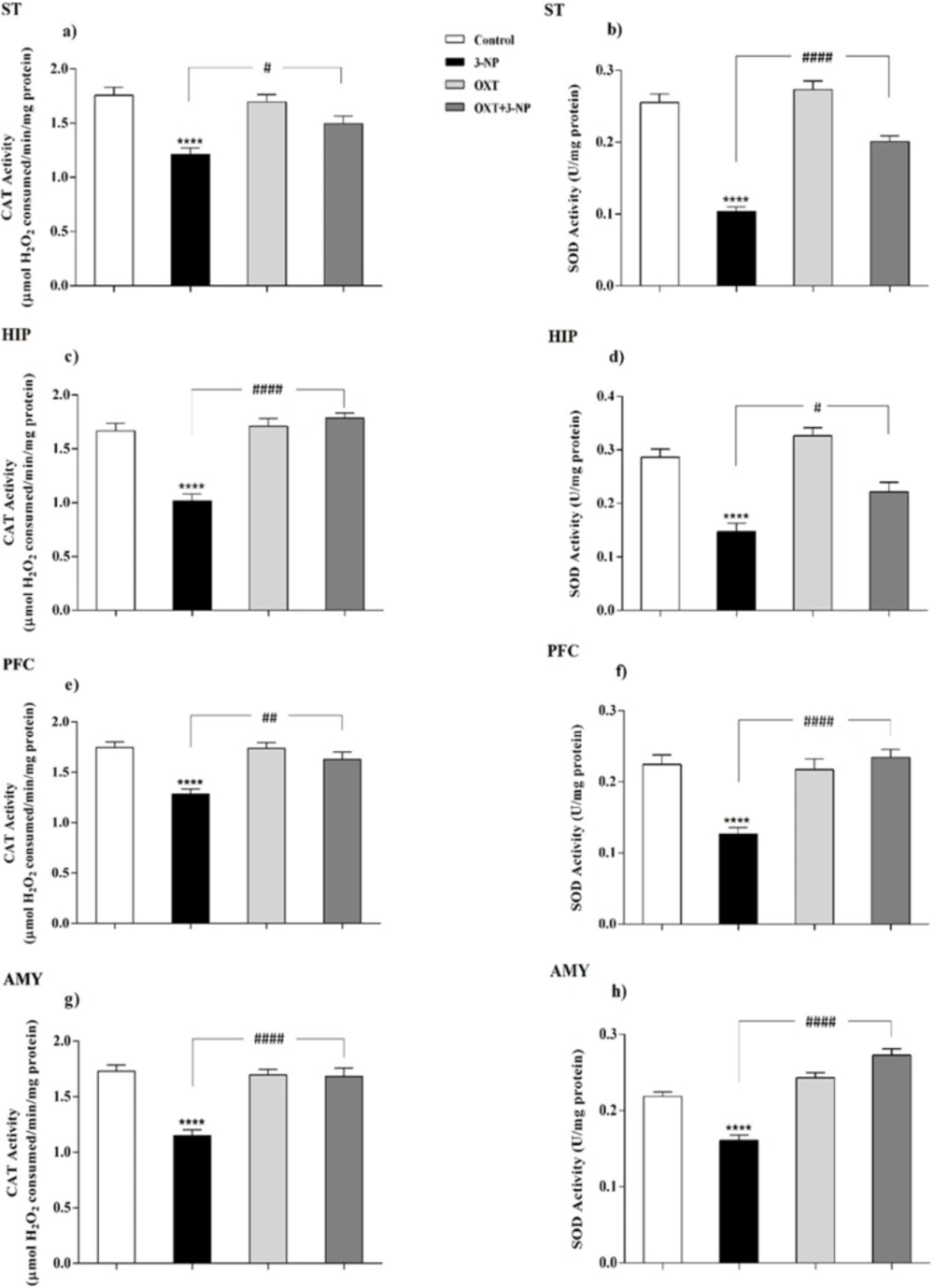
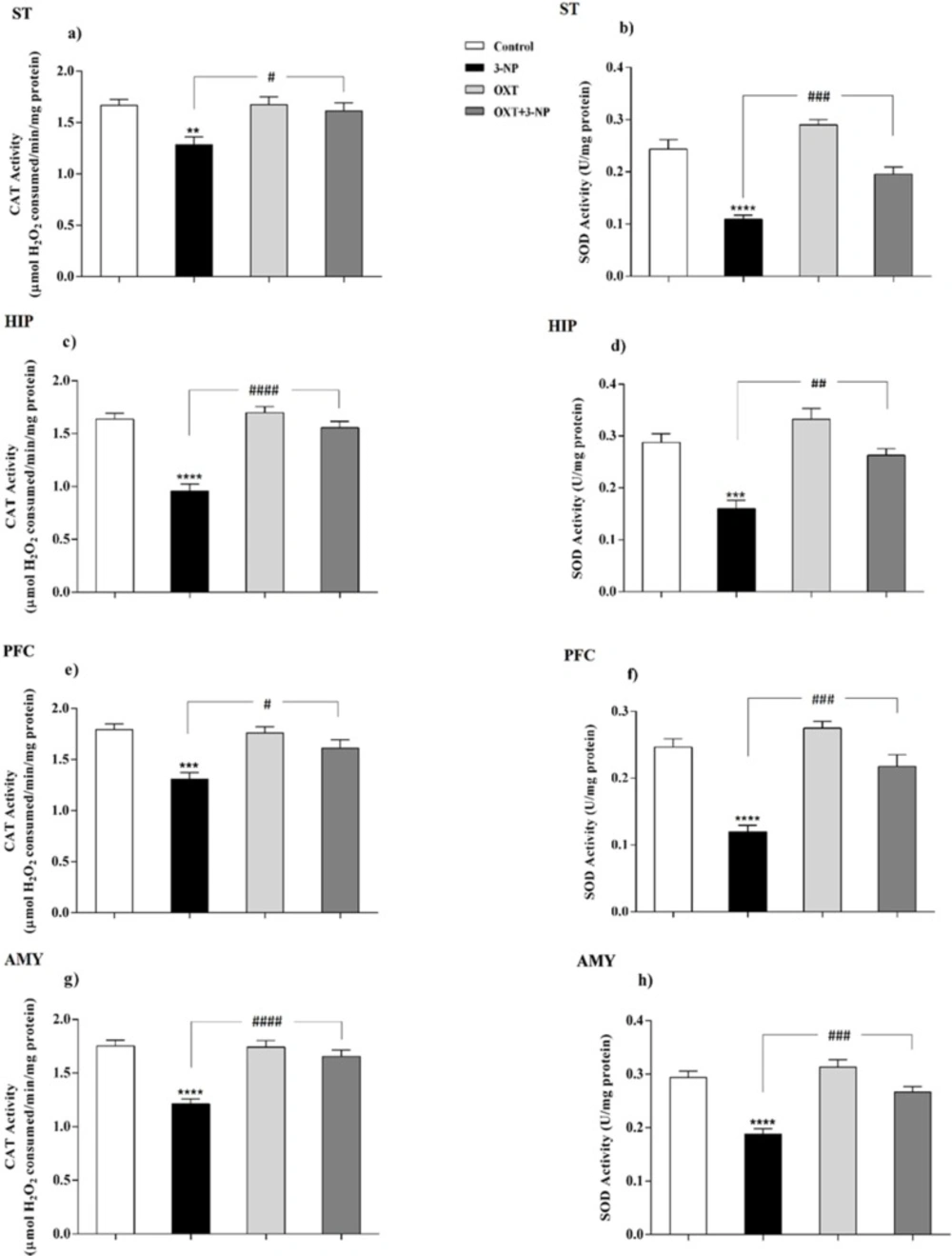
Peroxisomes are essential in cellular metabolism and their deficiency causes severe pathological situations, particularly in the brain and liver (
38). They play a safeguard role in protection against ROS, principally through having CAT, and also partly because of possessing SOD (
39). Although the mitochondrial matrix is the main place of SOD, it has been reported that peroxisomal membrane enjoys its presence as well (
39). However, there is no clear indication of the role of peroxisomes in neurodegenerative diseases such as HD and it is not still clear how metabolic dysfunction triggers these defects. So, it was tempting to consider its possible involvement. For this purpose, the distribution of peroxisomes was investigated by evaluating the amount of peroxisomal membrane proteins (PMP70, Pex14) along with assay of CAT activity as a peroxisomal matrix enzyme. Besides, regarding the role of peroxisome in the maintenance of redox hemostasis the activity of SOD was measured as an antioxidant enzyme.
3-NP diminishes oxidative phosphorylation in the mitochondrial respiratory chain, reduces the level of available ATP, causing metabolic inhibition and abnormal production of ROS (
40,
41). 3-NP could be used in laboratory animals to replicate the phenotype of HD associated with degeneration of the ST. Moreover, previous studies have illustrated that oxidative stress plays a fundamental role in the pathogenesis of HD (
18-
20). The results demonstrated that 3-NP injection reduced the activities of SOD and CAT in the ST of male rats which were in line with previous study (
42). In addition to the ST, 3-NP also causes metabolic dysfunction and neuronal degeneration in other brain regions therefore, other areas including PFC, HIP, and AMY were investigated and these destructive effects of 3-NP were also observed in other studied brain areas (
43). Peroxisomes are dynamic organelles, that environmental stimuli could affect dynamic regulation of their size, abundance, and function (
44-
46) and recently, a possible physical interaction of the Pex11 protein involved in the biogenesis of the peroxisomes with the mitochondrial Mdm34 protein has been suggested (
47,
48). Mitochondrial distribution and morphology protein 34 (Mdm34)-Pex11 interaction might have dynamic functions in the regulation of ROS homeostasis, metabolite exchange between the two organelles, or the modulation of respiratory efficiency (
49). Furthermore, the end product of β-oxidation in human peroxisomes, shortened fatty acyl-CoA, is shuttled into mitochondria to fuel the Krebs cycle and respiration and therefore it can be assumed that the mitochondrial respiratory chain extensively supply the ATP to peroxisomal α-oxidation pathway (
3-
5). As a result, the efficient function of peroxisome is necessary to avoid oxidative damage (
6). Our study showed that 3-NP injection significantly reduced the level of both Pex14 and PMP70 in different brain regions. Pex14 is a membrane-anchored peroxin, involved in the peroxisomal import of matrix proteins and considers as a marker for peroxisomal number (
36,
50 and
52). Recently it has been demonstrated that Pex14 has been participated in microtubule-based peroxisome motility (
53). Berger’s group demonstrated a correlation between increased peroxisomal volume density and impaired peroxisome trafficking in neurons of human AD brain, associated with (
54). With this in mind, our study demonstrated that 3-NP injection decreased the level of this protein and impaired peroxisomes function. This likely explains that peroxisomal trafficking dysfunction may contribute to the 3-NP-induced neurotoxicity. PMP70, as a major component of mammalian peroxisomal membranes, is now accepted as a good candidate for the overall size of peroxisomal population (
55). This ATP binding cassette transporter has the duty of transporting long and branched-chain fatty acyl-CoAs so is an important factor in metabolic function of peroxisomes (
37). Fanelli
et al. indicated the specific increase in early AD of PMP70 that could reflect the need for a more efficient acyl-CoA β-oxidation (
56). This increase is a compensatory mechanism in response to Aβ-induced mitochondrial dysfunction and energy metabolism (
57,
58). They showed that in transgenic mice of ages between 3 and 6 months, when several hallmarks of AD pathology appeared, PMP70 is significantly decreased that indicating decreased efficiency of peroxisomal β-oxidation. In this regard, our study showed that after the development of symptoms in 3-NP-induced HD-like model, the expression level of PMP70 decreased suggesting that mitochondrial dysfunction upon 3-NP injection results in peroxisomal dysfunction and finally decreased PMP70 level. According to our results, OXT pretreatment in 3-NP-induced neural toxicity model, significantly increased the antioxidant capability through increasing the activity of the CAT and SOD enzymes. This effect appeared in different brain regions that was affected by 3-NP, which is in agreement with previous reports describing the antioxidant effects of OXT (
25-
27). In addition, it has been shown OXT increases glucose uptake in cardiomyocytes in critical situations such as hypoxia through calcium-calmodulin kinase (Ca-CAMKK) and AMP-activated protein kinase (AMPK) pathways (
59). AMPK activation promotes glucose uptake, glycolysis, limits apoptosis, and cell damage (
60,
61). Altogether, our findings suggest that OXT polished the function of PMP70 and Pex14 perhaps by improving mitochondrial function. This protective effect of OXT consequently, improves antioxidant system capability through enhancing the CAT and SOD activities that play central roles in the predominant antioxidant activity of peroxisomes and mitochondries in mammalian cells. Furthermore, it has been reported that OXT exerts its effects in a sex and context dependent manner (
62,
63). Also, it has been previously described that the male rats have a commonly higher rate of hepatic β-oxidation when compared with females (
64). Therefore, we asked whether the male and female rats respond differently to OXT. Our results showed no sex-dependent effect in the presence of OXT. However, many studies have addressed its sex-related effects. We do not have a clear-cut explanation for these inconsistent observations, but it could be hypothesized that the context-dependency effects of OXT is an important factor in its sex-dependent effects. Secondly, most of the measured factors in this study are metabolism-related, in a way that their abundance is strongly dependent on cellular condition and variation in lipid metabolism (
3,
5). So it seems that aforementioned question could be answered better with investigating some other structural proteins in peroxisome and of course with choosing different contexts for male and female rats. This work is now in progress in our lab.