In this study, we assessed the effects of VPA, a histone deacetylase inhibitor, to determine its effects on the expression of genes involved in tumor metastasis in the A549 lung cancer cell line. Histone acetylation is one of the most important epigenetic mechanisms modulating gene expression. We observed here that VPA decreased the viability of A549 cells upon 72 h incubation in a dose-dependent manner. Inline, a dose-dependent increase was observed at both gene and protein levels of Nm23-H1, a tumor suppressor in the cancerous cells. On the other hand, the expression of metastatic tumor indicators, CD44v6, MMP-2, and MMP-9 decreased in A549 cells exposed to VPA. These changes further were accompanied by increased caspase-3 activity in VPA treated A549 cells.
Nm23-H1 is a multifunctional protein with nucleoside diphosphate kinase, histidine kinase, and DNase activities (
16). Studies have shown the antineoplastic activity of HDACs in hematologic and solid malignancies. Scientific evidence also supports the key role of HDACs in down-regulating of genes involved in tumor metastasis and invasion both
in-vitro and
in-vivo (
17,
18). HDAC inhibitors can suppress tumorigenesis by halting cancer cells migration, invasion, and growth, as well as by inducing apoptosis (
19,
20). In this study, we showed that VPA as an HDAC inhibitor, induced the expression of
Nm23-H1, a tumor suppressor downregulated in highly metastatic cancers (
4).
Nm23-H1 upregulation can induce DNA damage and subsequently genomic instability (
21). Based on our findings and the effects of VPA on
Nm23-H1, epigenetic alternations of this gene in A459 cancer cells may also be a mechanism involved in the metastatic behavior of lung cancer cells. Therefore, applying HDACs inhibitors, particularly VPA, may provide a therapeutic option in this type of cancer.
HDACs also can induce apoptosis in cancer cells (
19,
20). Accordingly, it has been shown that VPA-induced apoptosis may involve an increase in acetylation of histones and tubulin, a study conducted in a gastric cancer cell line (
22). In our study, the apoptotic effect of VPA on the lung cancer cell line A549 was accompanied by an increase in caspase-3 activity. This result agreed with a previous study that reported the up-regulatory effect of VPA on caspase-3 (
23).
CD44 is a cell-surface glycoprotein involved in cell–cell and cell–matrix adhesion, cell migration, and metastasis. Among all
CD44 isoforms,
CD44v6 harboring a mutation in exon 11 plays an important role in enhancing the adhesive ability of tumor cells (
24). The adhesion of cancer cells to the basal epidermal components such as collagen, integrin, and fibronectin is mediated by
CD44v6. An evidence-based report showed that an increase in the expression of
CD44v6 altered the physicochemical properties of tumor cells and increased their metastatic potential (
25).
CD44v6 the We showed the inhibitory effects of VPA on
CD44v6 at both gene and protein levels in the A459 cell line in the present study. This observation indicates a potential inhibitory impact for VPA on tumorigenesis in lung cancer.
Our results also revealed an inhibitory effect for VPA on the expression of
MMP-2 and
MMP-9 genes in the A459 cell. Type I collagenases such as
MMP-2 and
MMP-9 participate in cancer growth and invasion by degrading extracellular matrix (ECM) (
26).
MMP-2 and
MMP-9 are zinc-dependent ECM degrading enzymes involved in the metastatic activity of tumor cells (
27). According to the effects of VPA on the expression of these genes in A459 cells observed here, HDACs and epigenetic mechanisms may be involved in lung cancer progression. Therefore, HDACs inhibitors such as VPA can provide a viable therapeutic agent in these cancers.
In the present study, the up-regulation of
Nm23-H1 upregulation was seen in concomitant with the down-regulation of
MMP-9. This phenomenon can promote a potent anti-metastatic effect on cancer cells. The interaction between
Nm23-H1 and
MMP-9 is controversial.
Nm23-H1 has been shown to increase
MMP-9 gene expression and its gelatinolytic activity (
28). In another report, however,
Nm23-H1 did not modify
MMP-9 expression (
29). Moreover,
Nm23-H1 upregulation has been related to
MMP-9 suppression in yet another report (
30). In our study, increased
Nm23-H1 expression was seen in parallel to decreased expression of
CD44v6,
MMP-2, and
MMP-9. This event, along with elevated caspase-3 activity, can finally result in apoptosis in A549 human lung cancer cells.
In conclusion, our study demonstrated the antitumor activity of VPA against the A549 lung cancer cell line. One possible mechanism may be the Inhibition of HDAC, which increased Nm23-H1 expression. Furthermore, VPA-treated A549 cells showed decreased expression of CD44v6, MMP-2, and MMP-9 considered as metastasis indicators in cancer. The underlying inhibitory mechanisms of VPA on tumor cells and its potential therapeutic role in cancer are yet to be investigated.
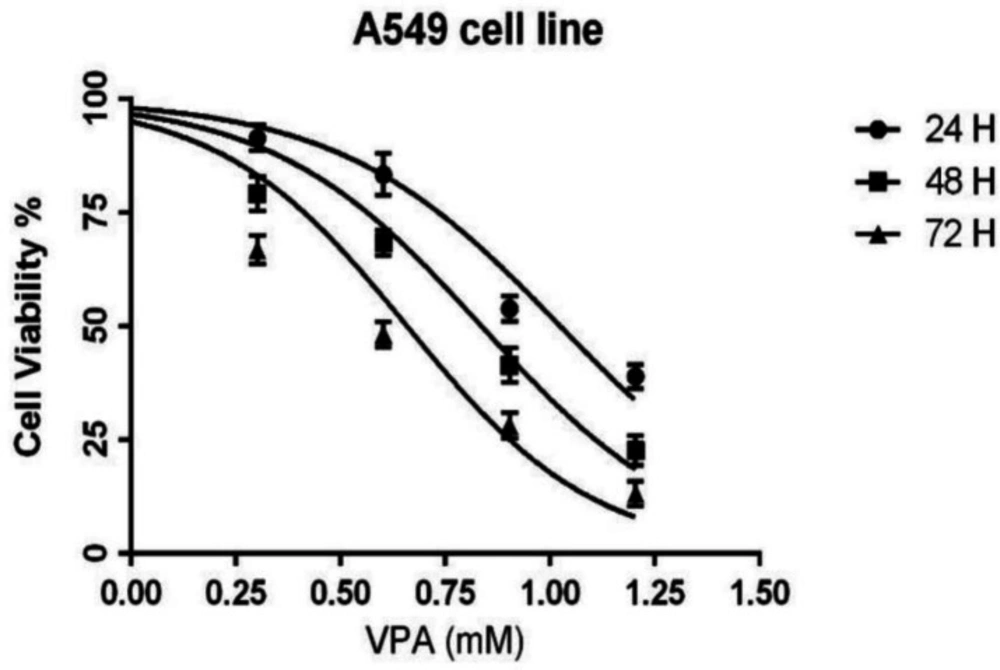
The effect of VPA (0-16 Mm) on the cell viability of A549 cells after 24, 48 and 72 h incubation. Results are expressed as means ± SEM, n = 3
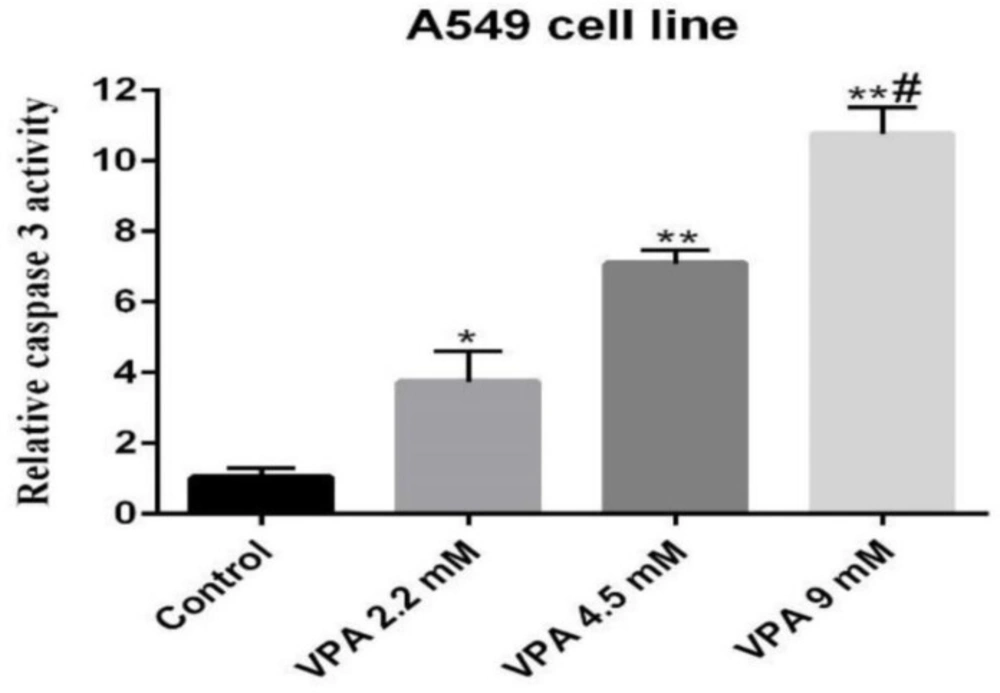
Relative caspase 3 activity was determined in 549 cell line treated with 2.2, 4.5 and 9 mM of VPA for 72 h. *(P ˂ 0.01), **(P ˂ 0.001) compared to control cells, #(P ˂ 0.001) compared to 2.2 and 4.5 mM of VPA
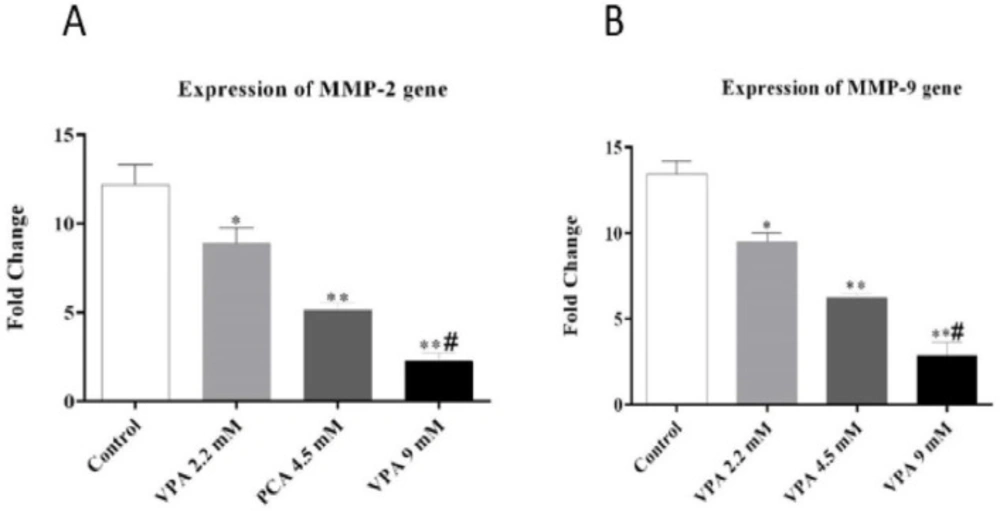
Effect of different concentrations of VPA on the expression of MMP-2 and MMP-9 gene after 72 h incubation. *(P ˂ 0.01), **(P ˂ 0.001) compared to control cells, #(P ˂ 0.001) compared to 2.2 and 4.5 mM of VPA
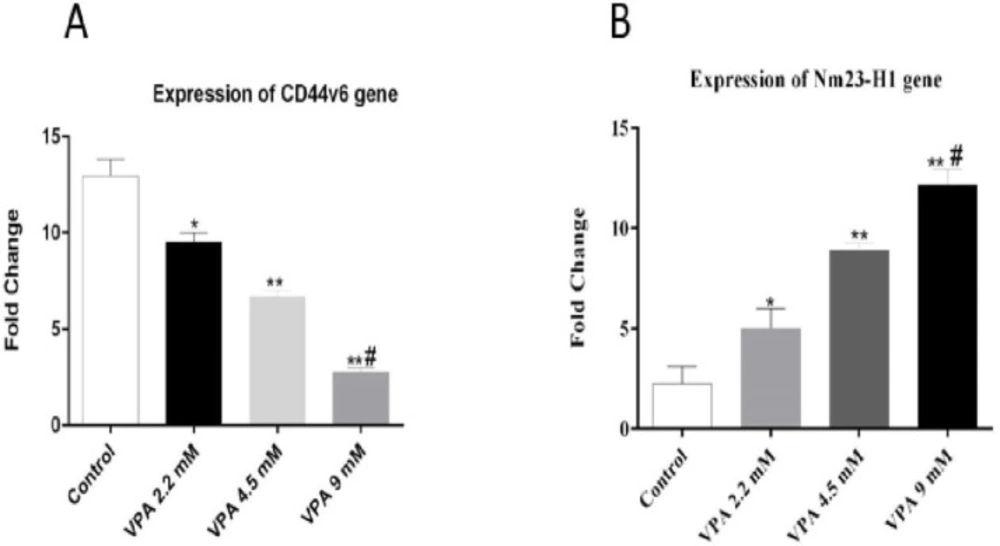
Effect of different concentrations of VPA on the expression of Nm23-H1 and CD44v6 gene after 72 h incubation. *(P ˂ 0.01), **(P ˂ 0.001), compared to control cells, #(P ˂ 0.001) compared to 2.2 and 4.5 mM of VPA
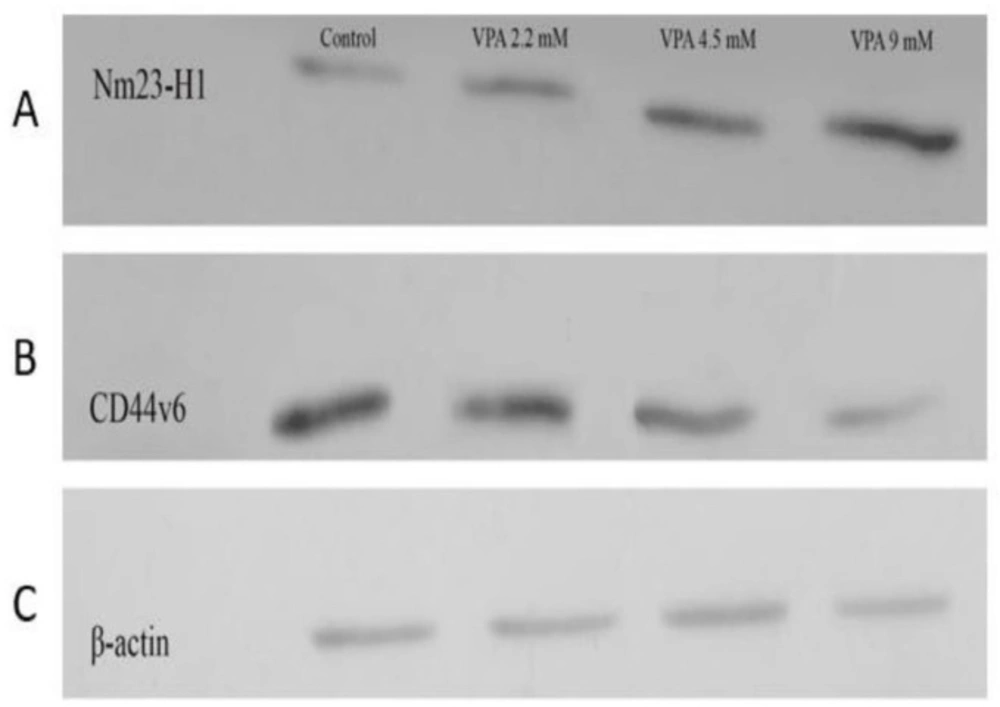
Effect of VPA on the expression of Nm23-H1 and CD44v6 protein in A549 cell line. VPA increased and reduced the expression of Nm23H1 and CD44v6 protein in A549 after 72 h, respectively
| Gene | F/R | Primer sequences (5'3') |
|---|
| Nm23H1 | Forward | TTAATCAGATGGTCGGGGAT |
| Reverse | GATCTATGAATGACAGGAGG |
| CD44V6 | Forward | GTCGATGCTAGCTAGCCGTAGCATG |
| Reverse | CGAGCTAGTCGTAGTCGATCGATCG |
| MMP2 | Forward | TCTCCTGACATTGACCTTGGC |
| Reverse | CAAGGTGCTGGCTGAGTAGATC |
| MMP9 | Forward | CCTTGTGCTCTTCCCTGGAG |
| Reverse | GGCCCCAGAGATTTCGACTC |
| GAPDH | Forward | AATCCCATCACCATCTTCCA |
| Reverse | TGGACTCCACGACGTACTCA |