
Introduction
Experimental
| Primer names | Primer sequences from 5' to 3' direction | PCR product length |
|---|---|---|
| IntI forward | GCCTTGCTGTTCTTCTACGG | |
| 558 bp | ||
| IntI Reverse | GATGCCTGCTTGTTCTACGG |
| Primer names | Primer sequences from 5' to 3' direction | Name of target gene | PCR product length |
|---|---|---|---|
| VEB-1A Forward | CGACTTCCATTTCCCGATGC | blaVEB | 643 bp |
| VEB-1B Reverse | GGACTCTGCAACAAATACGC | ||
| GES-1AForward | ATGCGCTTCATTCACGCAC | ||
| GES-1B Reverse | CTATTTGTCCGTGCTAAGG | blaGES | 860 bp |
| Primer names | Primer sequences from 5' to 3' direction | PCR product length |
|---|---|---|
| ERIC forward | CACTTAGGGGTCCTCGAATGTA | |
| 558 bp | ||
| ERIC Reverse | AAGTAAGTGACTGGGGTGAGCG |
| Primer names | Primer sequences from 5' to 3' direction | Name of target gene | PCR product length |
|---|---|---|---|
| ExoS Forward | CGTATGAGTCAGCAAGGGCG | ||
| ExoS | 118 bp | ||
| ExoS Reverse | GCGATGTGGTCACTGGCTTC | ||
| ETA Forward | GACAACGCCCTCAGCATCACCAGC | ||
| ETA | 396 bp | ||
| ETA Reverse | CGCTGGCCCATTCGCTCCAGCGCT | ||
| oprL Forward | ATGGAAATGCTGAAATTCGGC | oprL | |
| 504 bp | |||
| oprL Reverse | CTTCTTCAGCTCGACGCGACG | oprL | |
| oprI Forward | ATGAACAACGTTCTGAAATTCTCTGCT | oprI | |
| 249 bp | |||
| oprI Reverse | CTTGCGGCTGGCTTTTTCCAG | oprI | |
| Alg Forward | TTCCCTCGCAGAGAAAACATC | Alg | |
| 520 bp | |||
| Alg Reverse | CCTGGTTGATCAGGTCGATCT | Alg |
| Primer sequences from 5' to 3' direction | Name of target gene | PCR product length |
|---|---|---|
| AATCGCCGTCCAACTGCATGCG | Exo T | |
| 152 bp | ||
| TGTTCGCCGAGGTACTGCTC | Exo T | |
| CCGTTGTGGTGCCGTTGAAG | Exo U | |
| 134 bp | ||
| CCAGATGTTCACCGACTCGC | Exo U | |
| CGGATTCTATGGCAGGGAGG | Exo Y | |
| 289 bp | ||
| GCCCTTGATGCACTCGACCA | Exo Y |
| Number of strains in each group | Group name |
|---|---|
| 3 | A |
| 1 | B |
| 2 | C |
| 2 | D |
| 1 | E |
| 1 | F |
| 2 | G |
| 2 | H |
| 1 | I |
| 5 | J |
| 4 | K |
| 2 | L |
| 8 | M |
| 1 | N |
| 5 | O |
| 2 | P |
| 1 | Q |
| 2 | R |
| 1 | S |
| 2 | T |
| 1 | U |
| 1 | Y |
| Number of strains in each group | Group name |
|---|---|
| 1 | A |
| 2 | B |
| 1 | C |
| 1 | D |
| 2 | E |
| 1 | F |
| 2 | G |
| 3 | H |
| 1 | I |
| 4 | J |
| 4 | K |
| 2 | L |
| 1 | M |
| 1 | N |
| 8 | O |
| 14 | P |
| 2 | Q |
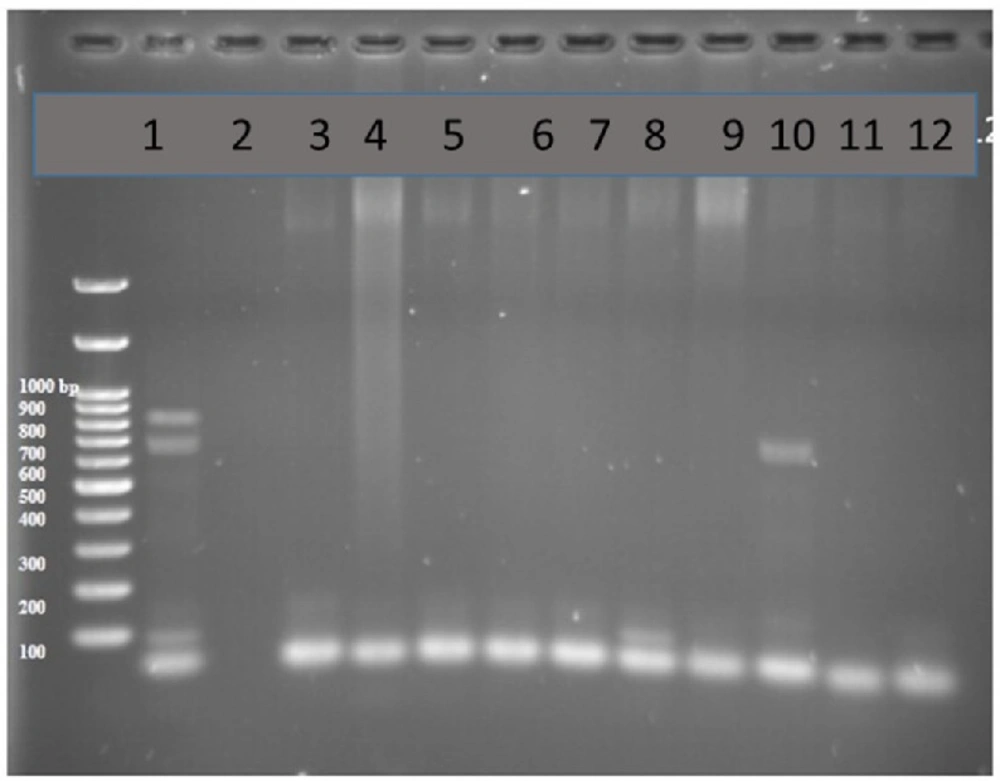
PCR products in well 1 represents two lengths of 680 bp and 643 bp as positive controls. Well number 2 represents negative control. Well number 10 contains PCR product of positive VEB encoding gene as DNA template
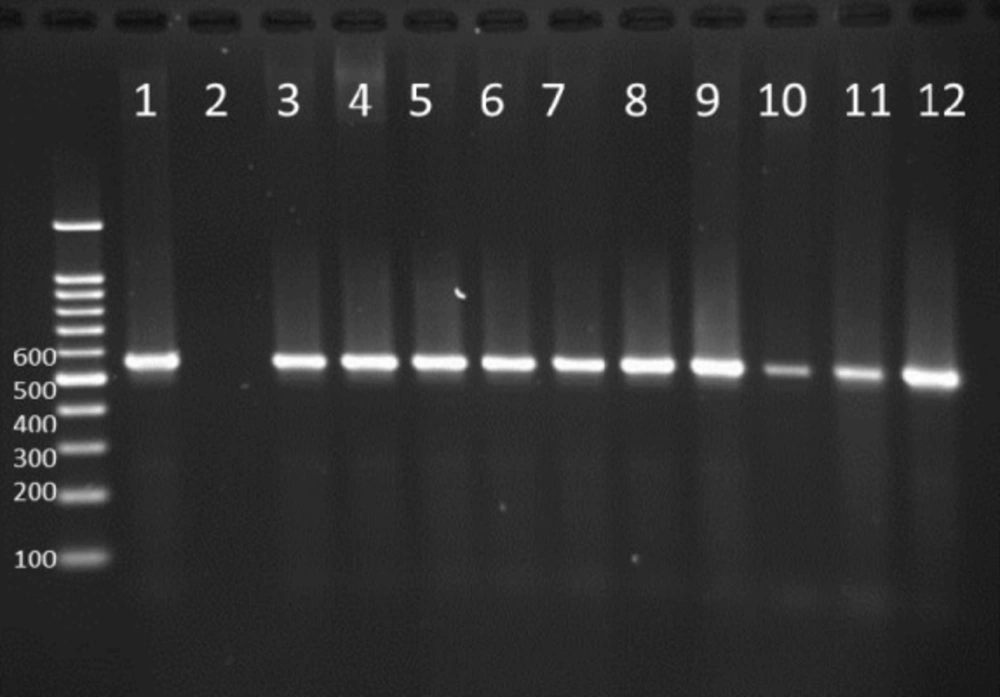
PCR products in well 1 represents positive control. Well 2 is negative control. Wells from number 3 to 12 contain PCR products of positive Integron I encoding gene as DNA template
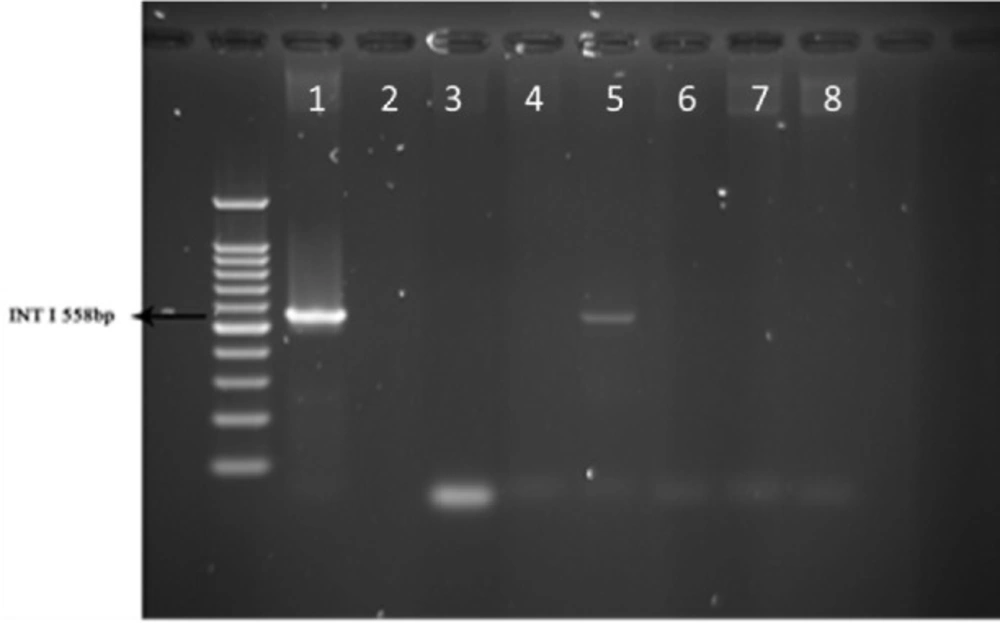
PCR products of well 1 represent positive control. The second well contains the PCR product of distilled water as template in PCR reaction as negative control. Wells from number 3 to 8 contain PCR products of animal samples amplified by INt I primers, well number 5 includes PCR product of Integron I encoding gene as DNA template
ERIC-PCR products of P. aeruginosa strains from human samples
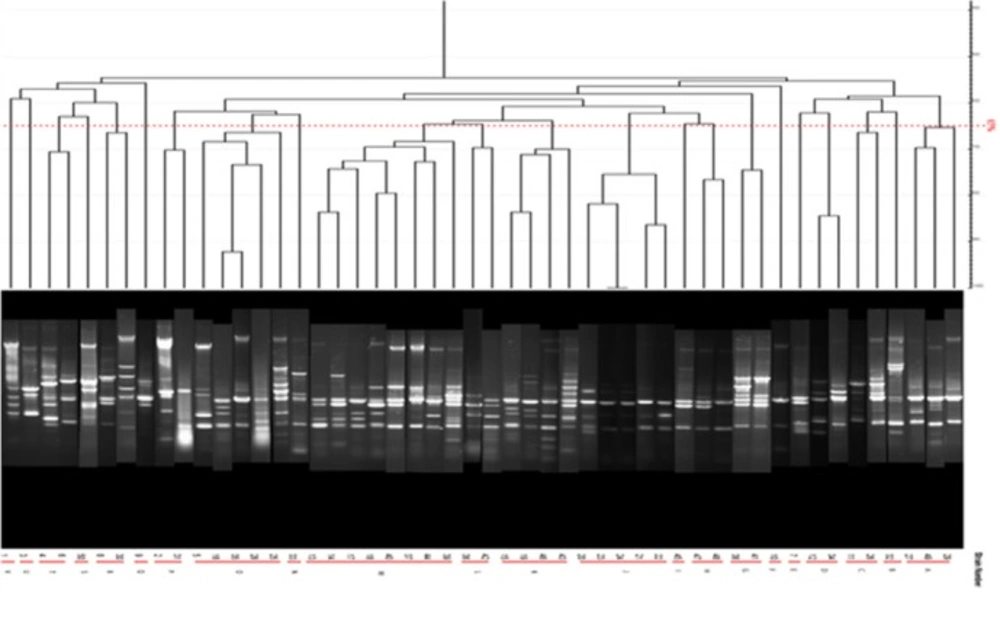
Dendrogram of the ERIC-PCR products of P. aeruginosa strains isolated from human samples are shown in Figure 5
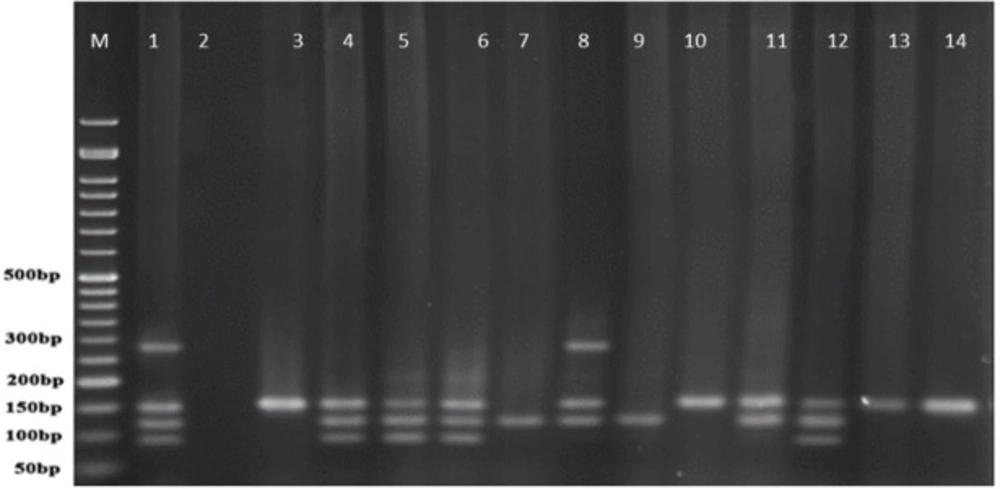
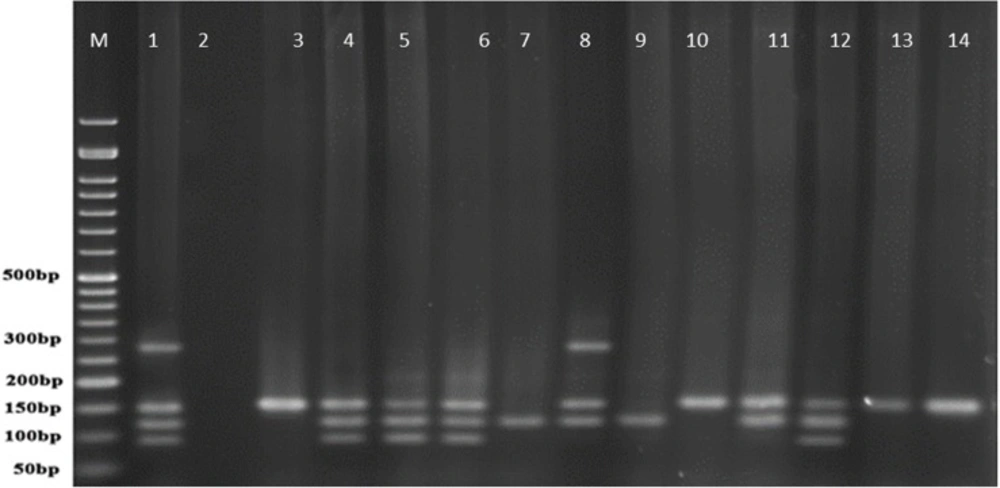
PCR products of well 1 represent positive control. The second well represents the PCR product of distilled water as template in PCR reaction as negative control. Wells from number 3 to 14 contain PCR products of human samples amplified with four specific primer pairs of exoenzymes under study
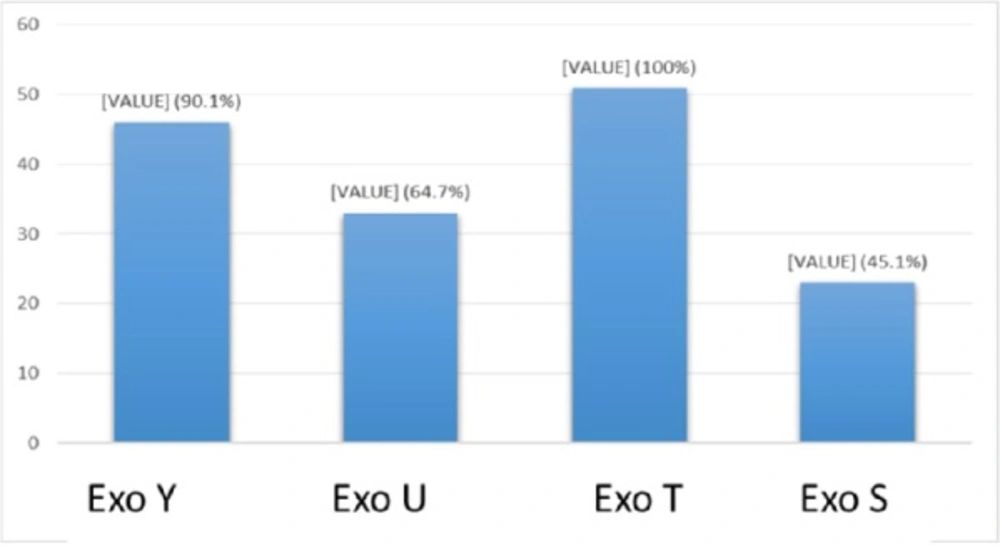
The frequency of exoenzymes Y, U, T, and S are shown in Graph 1. Exoenzyme T had the highest frequency of 100% among all studied samples taken from human patients
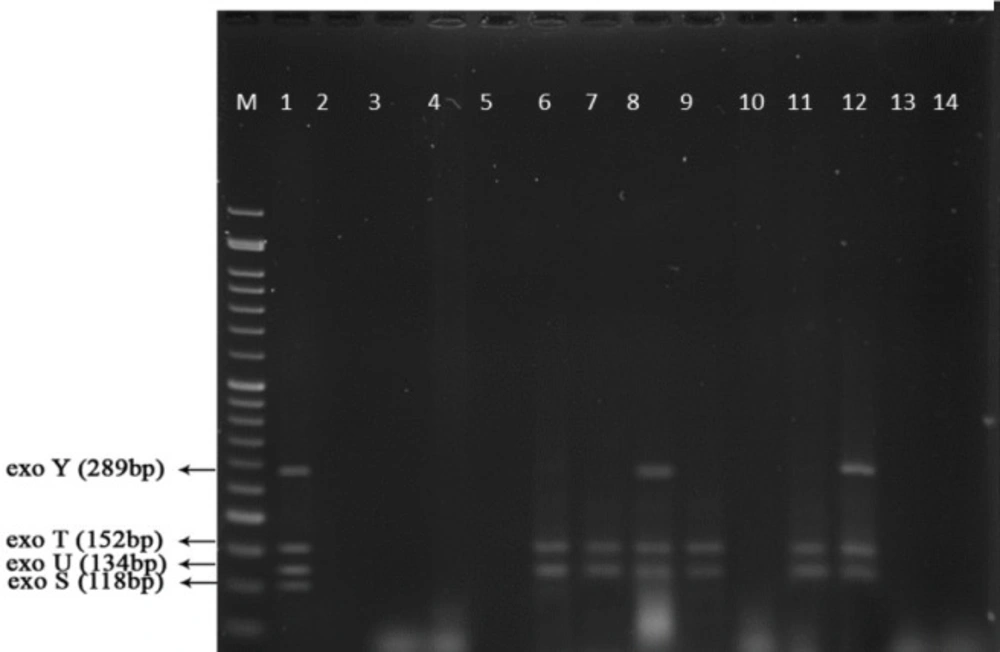
PCR products of well 1 represent positive control. The second well represents the PCR product of distilled water as template in PCR reaction as negative control. Wells from number 3 to 14 contain PCR products of human samples which are obtained from studying the exoenzymes
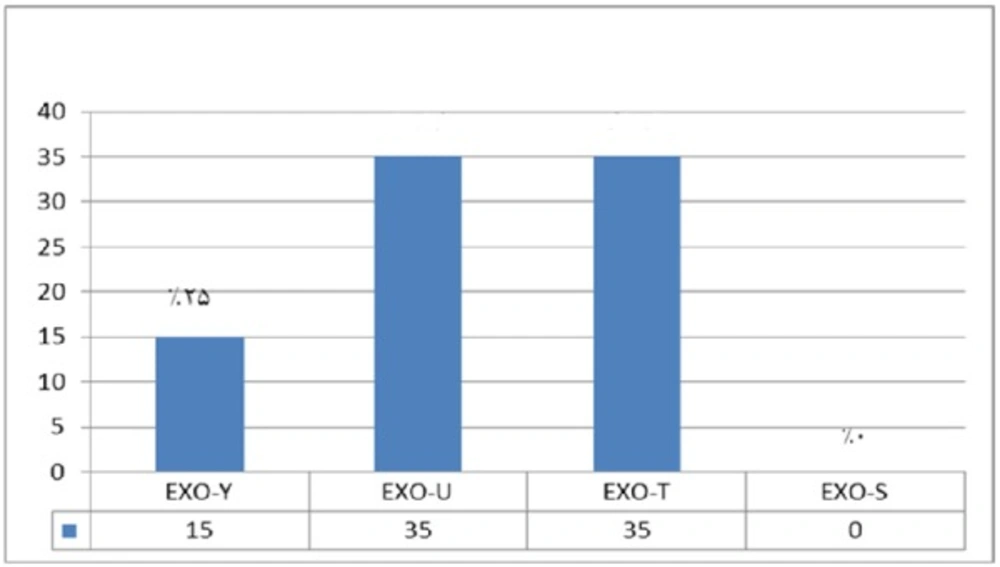
The frequency of exoenzymes Y, U, T and S are shown in Graph 1. Exoenzymes T and U had the highest frequency of 68.8% among all studied samples taken from animal isolated strains
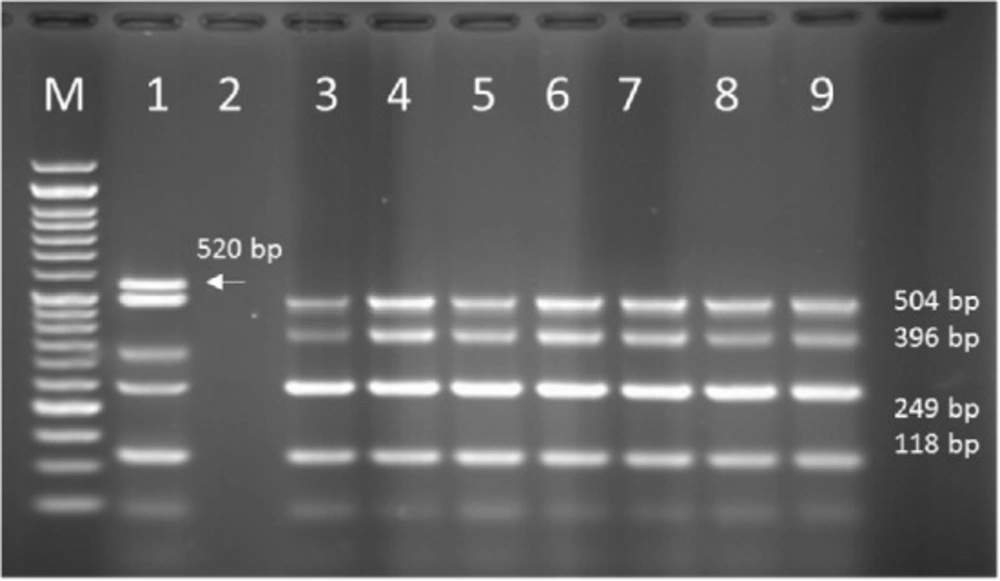
PCR products of Multiplex PCR results on DNA isolated from human on 1.5 % agarose gel electrophoresis showed specific bands of OPRL (504bp), OPRI (249 bp), ALGD (520 bp), Exo-S (118 bp), and EXT-A (396 bp) in comparison with DNA ladder.
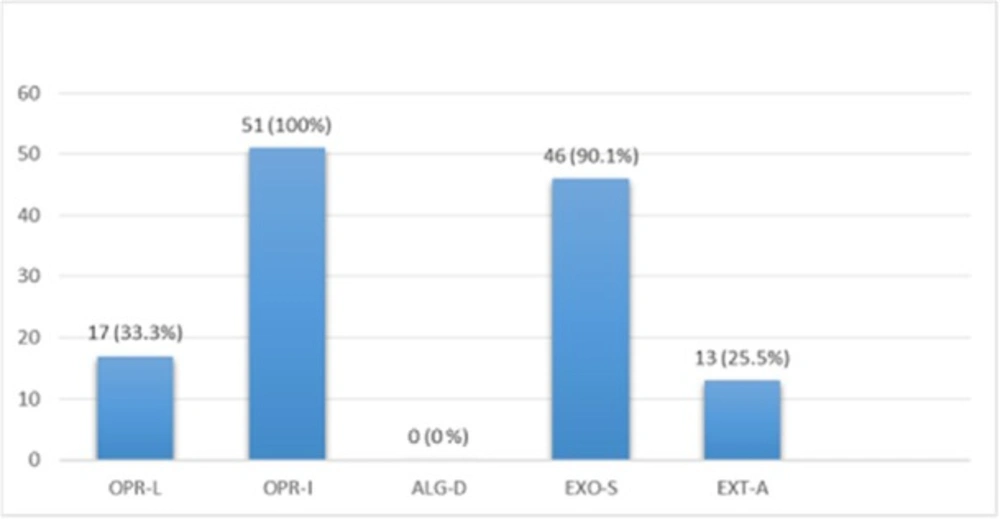
Multiplex-PCR results on strains isolated from human for detection of OPRL, OPRI, EXO-S, EXT-A, and ALGD encoding genes. ALGD and OPRI had the lowest and highest frequencies of 0% and 100% among isolated samples, respectively
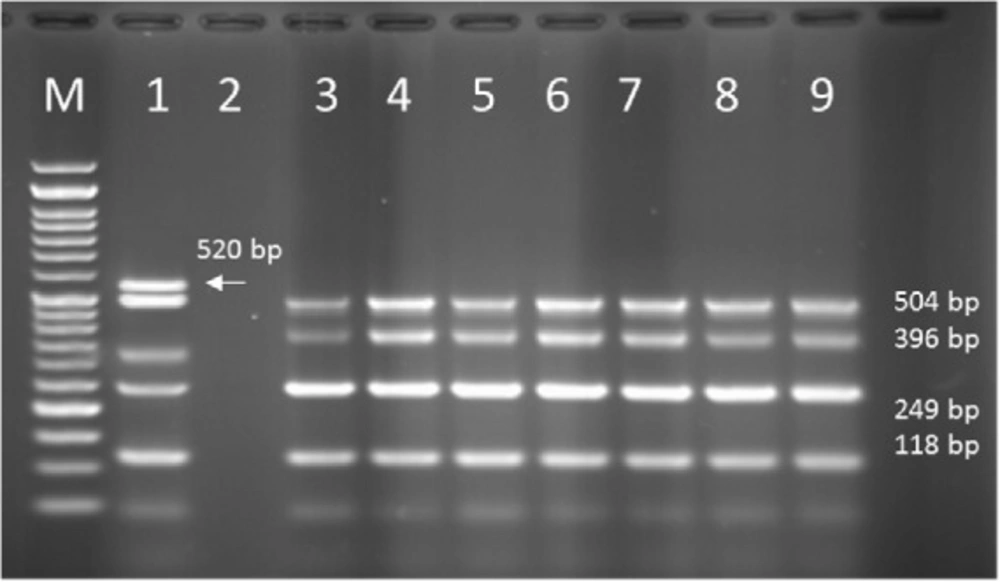
PCR products of Multiplex PCR results on DNA isolated from animal (cow) raw milk on 1.5 % agarose gel electrophoresis showing specific bands of OPRL (504 bp), OPRI (249 bp), ALGD (520 bp), Exo S (118 bp), and ext A (396 bp) in comparison with DNA ladder
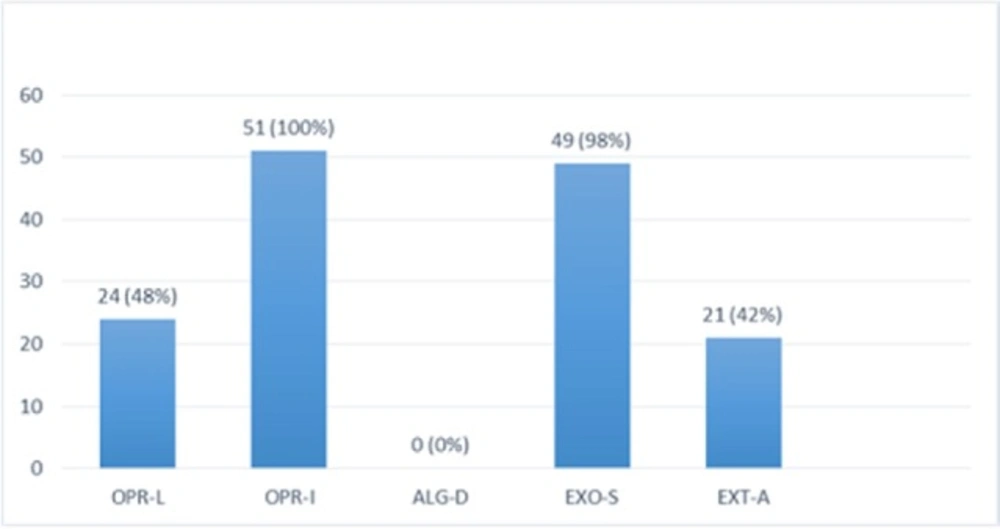
Multiplex-PCR results on strains isolated from cow raw milk for detection of OprL, OprI, exo S, ext A, and algD encoding genes; algD and OPRI have the lowest and highest frequencies of 0% and 100% among isolated samples, respectively
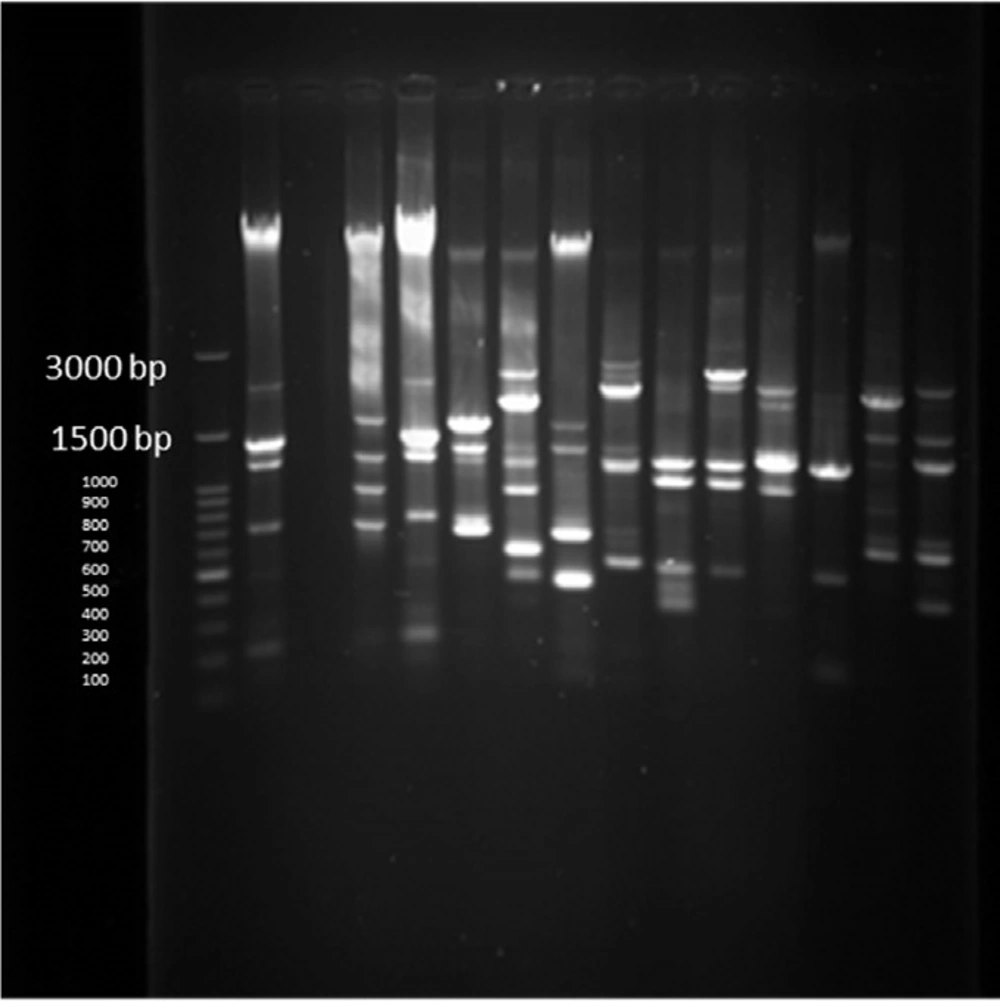
PFGE test results on 14 human samples (11 integron I positive and 1 Integron negative samples) and one Integron I positive and 1 Integron I negative samples from animals




