
Introduction
Experimental
Results
Discussion
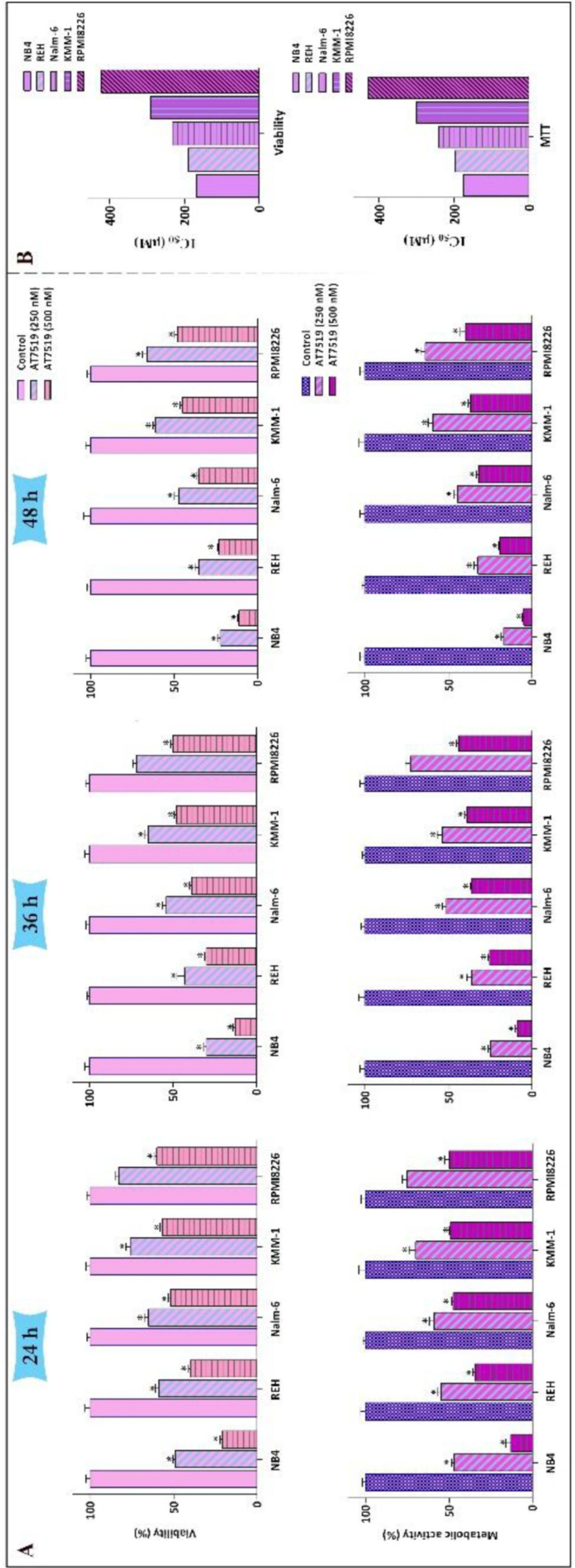
The anti-survival impact of AT7519 on a panel of leukemic cells. (A) Treatment of the cells with various concentrations of AT7519 abated both the viability and the metabolic activity in time- and concentration-dependent manners. (B) IC50 values were measured for two varied methods of survival assessment. Values are provided as mean ± SD of three separated tests. *P ≤ 0.05 represents considerable alters from untreated control
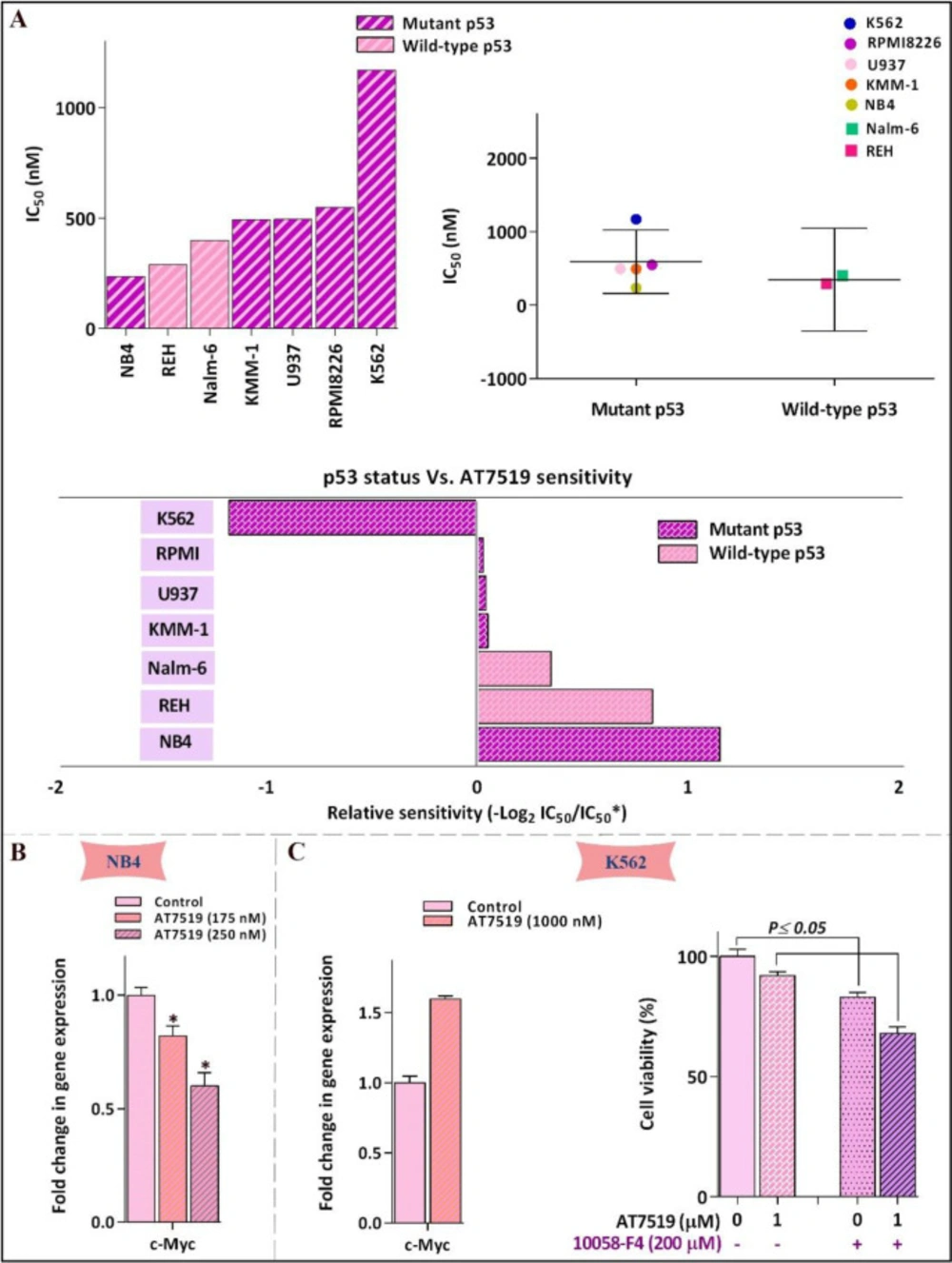
Leukemic cell response to AT7519 was irrespective of p53 status. (A) Relative sensitivity of all tested cell lines to AT7519. IC50 value of different leukemic cell lines to the inhibitor was determined using the formula: −log 2 (IC50 individual cell line/mean IC50 of all cells). There was no evidence of linkage between p53 status and cells sensitivity to AT7519. (B) AT7519 reduced the c-Myc expression in NB4 (C) but not c- in K562, as the less sensitive cell line. Values are provided as mean ± SD of three separated tests. *P ≤ 0.05 represents considerable alters from untreated control
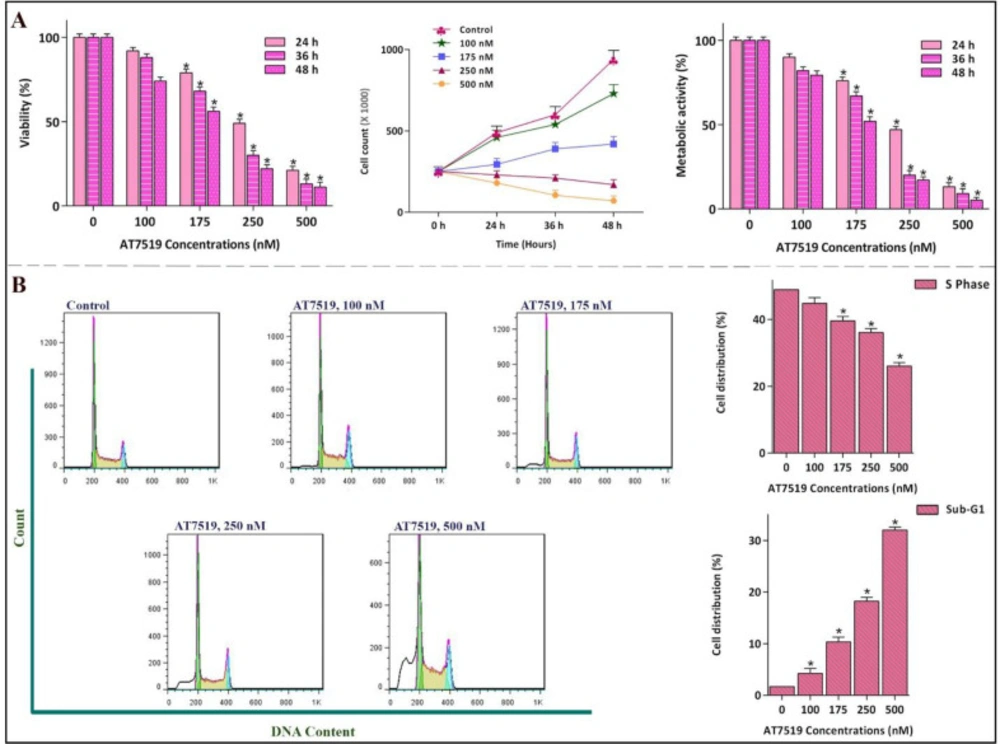
Anti-proliferative effect of AT7519 on NB4 and its impact on cell cycle progression. (A) the hindrance effect of AT7519 on CDKs was associated with a notable abatement in the viability and the number of viable cells in a concentration-dependent manner. (B) While the percentage of cells in S phase was lessened, cell population in Sub-G1 was escalated. Values are provided as mean ± SD of three separated tests. *P ≤ 0.05 represents considerable alters from untreated control
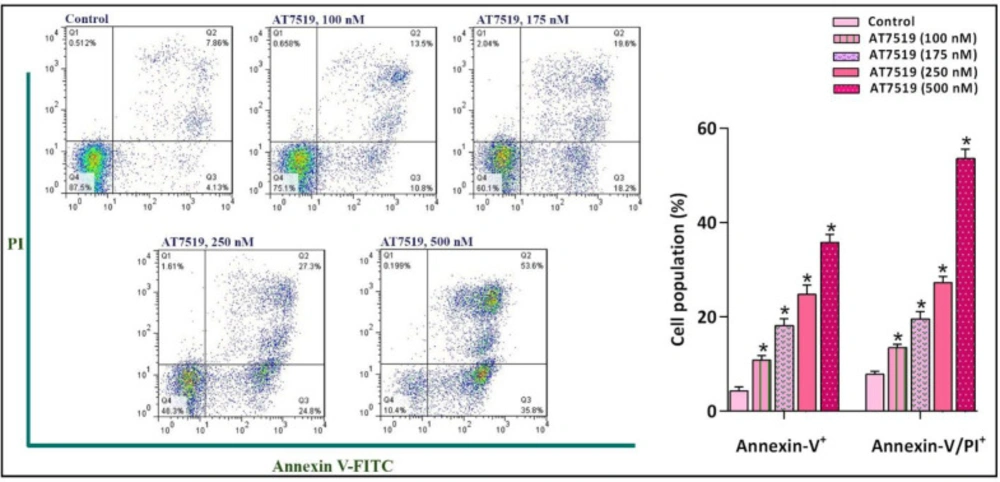
The suppression of CDK in NB4 cells was coupled with the induction of apoptotic cell death. FACS analysis of annexin-V/PI demonstrated that the inhibition of CDK using different concentrations of the inhibitor increased the proportion of both early and late apoptotic cells. Values are provided as mean ± SD of three separated tests. *P ≤ 0.05 represents considerable alters from untreated control
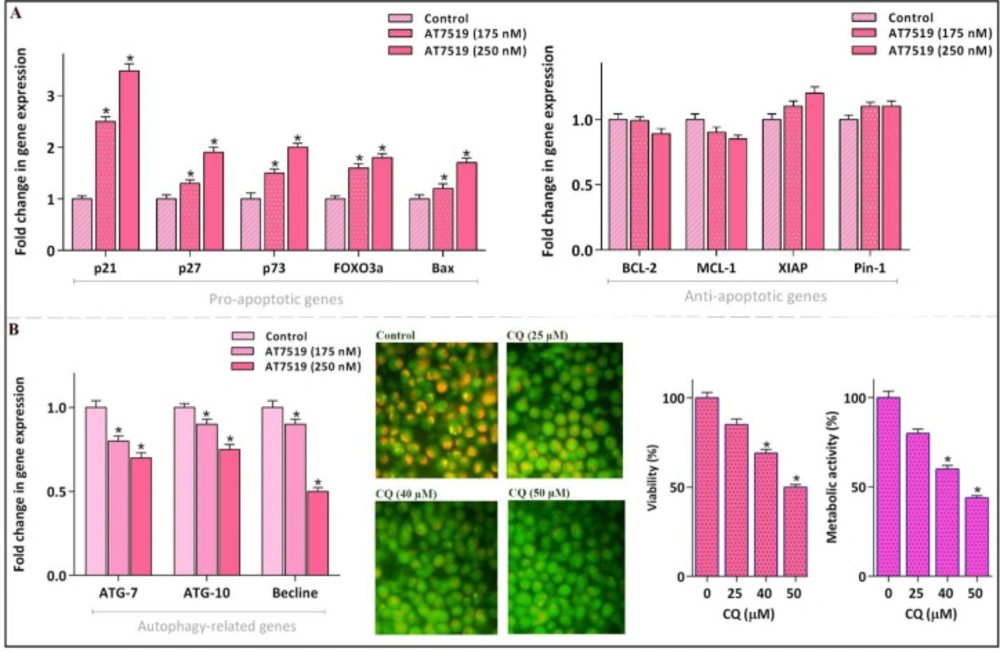
AT7519 could alter the apoptosis- and autophagy-related genes. (A) Although AT7519 up-regulated the mRNA expression levels of pro-apoptotic genes, it failed to significantly alter the transcription of the death repressor genes. (B) The inhibitor could remarkably reduce mRNA expressions of autophagy-related genes. The restriction of autophagy, as indicated by the decreased intensity of acridin orange, was coupled with the decreased survival of NB4 cell. Values are provided as mean ± SD of three separated tests. *P ≤ 0.05 represents considerable alters from untreated control
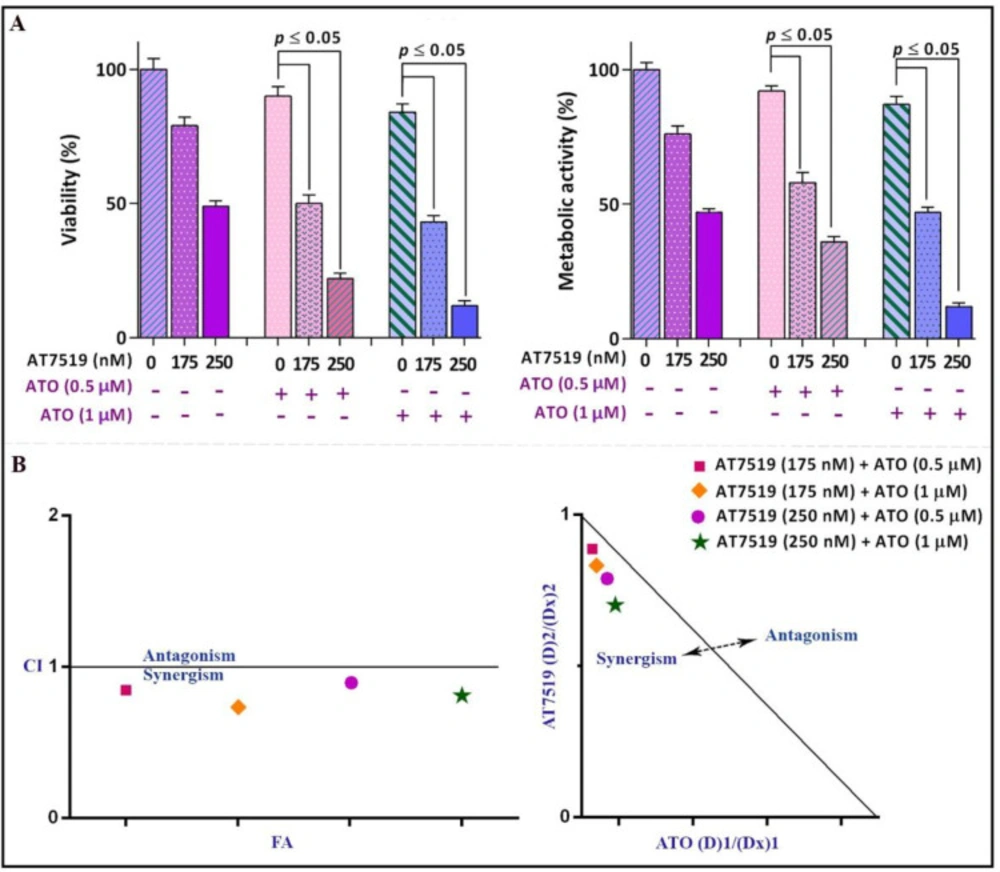
Surveying the synergistic impact of AT7519 and arsenic trioxide (ATO). (A) AT7519 could magnify the anti-leukemic influence of ATO in NB4 cells. (B) The results of both combination index (CI) and isobologram highlighted the synergistic effect between AT7519 and ATO. Values are provided as mean ± SD of three separated tests. P ≤ 0.05 represents considerable alters from untreated control
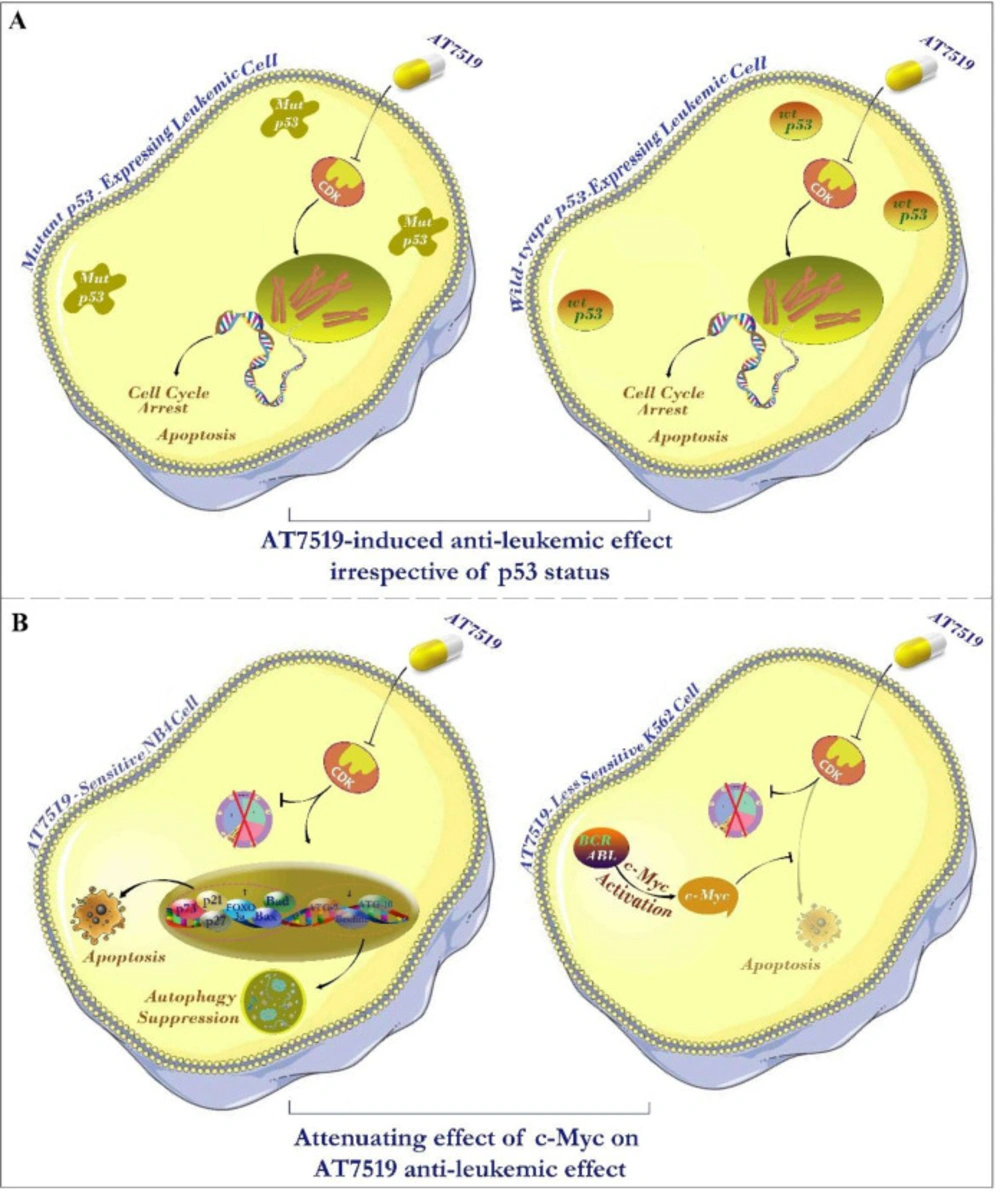
Schematic representation summarizes the resulting data of the present study. (A) Investigating the link between molecular status of p53 and leukemic cell sensitivity to the inhibitor hinted that AT7519 lessened the survival of leukemic cells independent of p53 status; suggesting the applicability of the inhibitor in both wild-type and deficient p53-expressing leukemic cells. (B) The eminent anti-tumor impact of this inhibitor on leukemic cells was associated with the induction of apoptotic cell death through switching the balance between pro- and anti-apoptotic genes. The effectiveness of AT7519 may be probably eclipsed by the compensatory activation of c-Myc in AT7519 less sensitive K562 cells, proposing that co-targeting both c-Myc and CDK would be a better strategy in chronic myeloid leukemia (CML).
