Human breast cancer is a complex illness caused by the joint interaction of genetic, epigenetic, and environmental factors (
23). Despite considerable developments in the treatment of breast cancer, current conventional therapies for this disease still have major limitations (
24). Thus, novel therapeutic strategies are needed to effectively modulate various carcinogenic mechanisms. Natural compounds such as diterpenes, play an important role in chemoprevention and in improving the effectiveness of cancer treatment by modifying the sensitivity of cancer cells to chemotherapeutics (
25).
Sclareol, as a diterpene, is a renowned phytochemical that was found to be able to potently reduce cell viability and induce apoptosis with an IC
50 at the micromolar level in estrogen receptor (ER)-positive MCF-7 cells (
26).
Compelling experimental evidence from both clinical trials and animal studies demonstrated that the JAK2/STAT3 pathway plays a pivotal role in the initiation, progression, and metastasis of breast cancer (
27,
28). The present study aimed to find out the molecular mechanism underlying sclareol anticancer activity, particularly in regards to its effects on the JAK2/STAT3 signaling pathway. In fact, on the basis of the existing evidence, we hypothesized that bioavailable sclareol alone, or in combination with cyclophosphamide may modulate breast carcinogenesis, primarily by inhibiting Stat3 activation.
Sclareol effects on cell viability and apoptosis
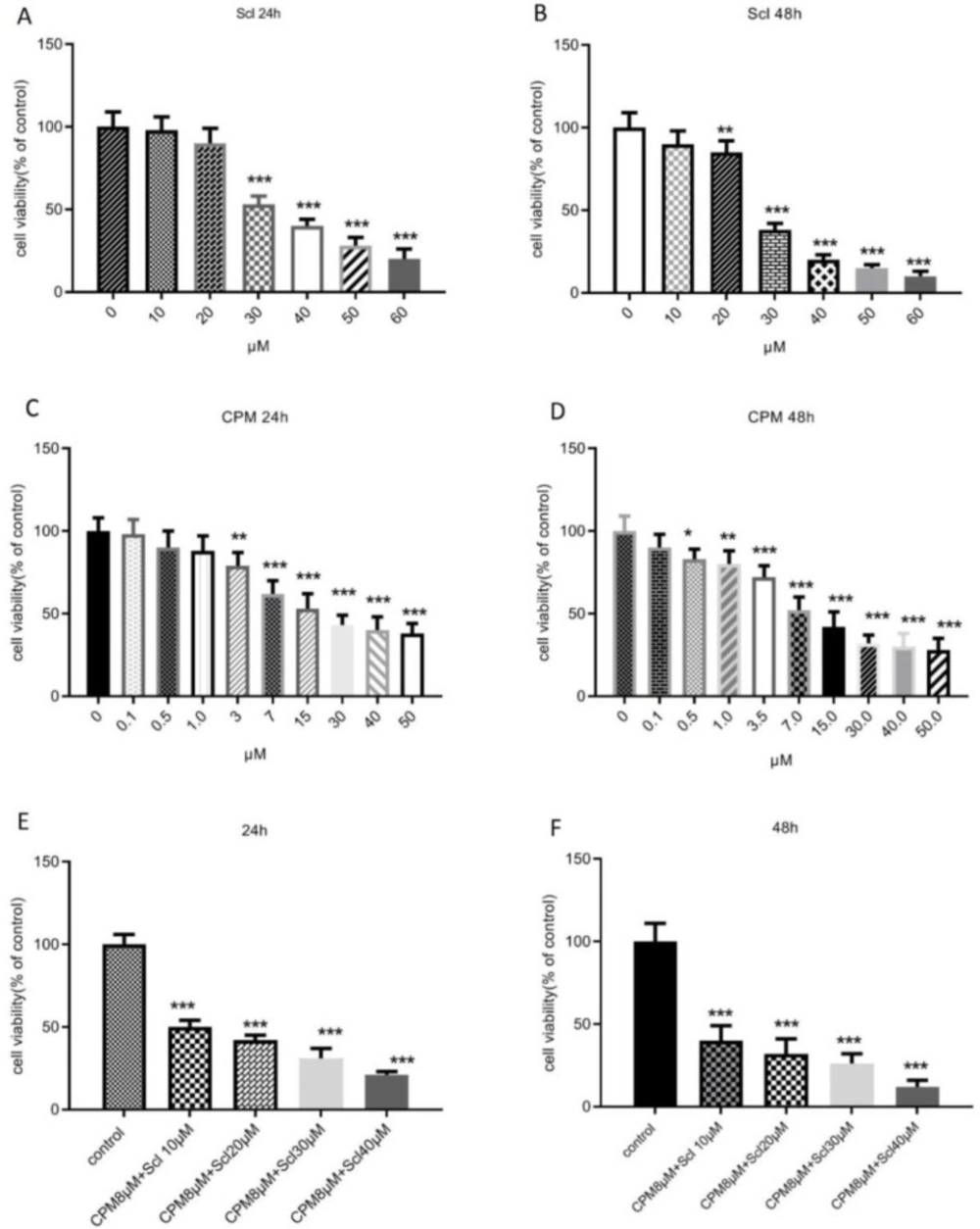
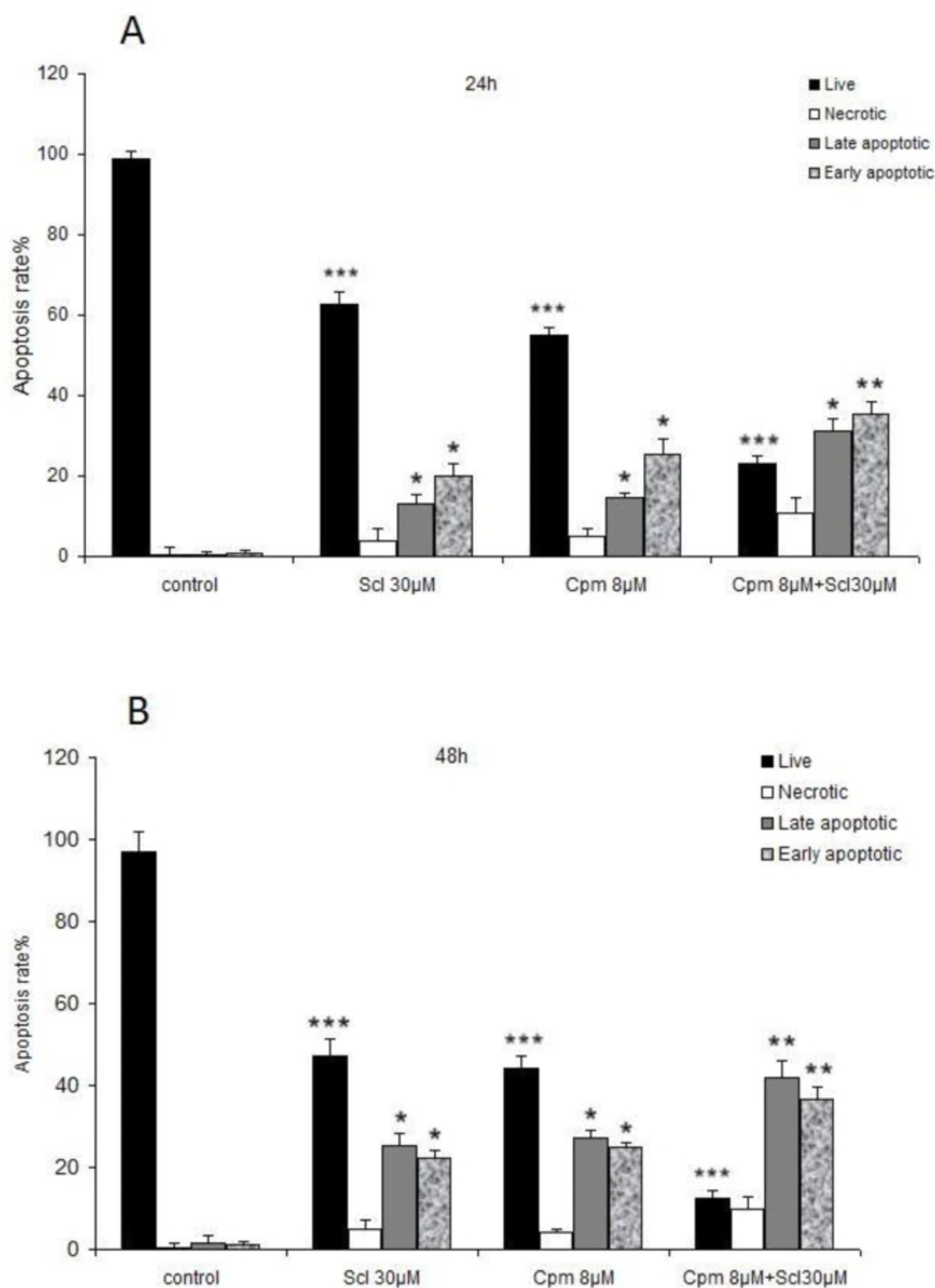
In this study, we showed that sclareol could effectively diminish cell viability and induce apoptosis in breast cancer cells at a concentration of 30 μM. In line with our findings, antiproliferative and apoptosis-inducing effects against breast cancer cells have been previously described for sclareol (
29). It has also been shown to reduce cell survival and induce cell death in other cancer types such as cervical cancer and osteosarcoma (
30,
31). Sclareol has been reported to sensitize cancer cells to various anticancer drugs. For example, sclareol enhances the efficacy of doxorubicin, etoposide, and cisplatin, against breast cancer cells (
29) and bortezomib in cervical cancer (
31). In the current study, we showed for the first time that co-administration of sclareol and cyclophosphamide exhibited a much more potent anti-proliferative effect against breast cancer cells and induced apoptosis more competently, suggesting that the combination of sclareol and cyclophosphamide might be considered as a suitable candidate for breast cancer treatment.
Apoptosis, the major type of programmed cell death, is an active cell suicide process. One of the major hallmarks of carcinogenesis is a circumvention of apoptosis by cancer cells. On the other hand, cancer cell death is an essential part of different strategies that exist for the elimination of malignant cells. Thus, initiation of cell death pathways as well as targeting the lesions that suppress cell death are exciting remedies for cancer treatment (
32).
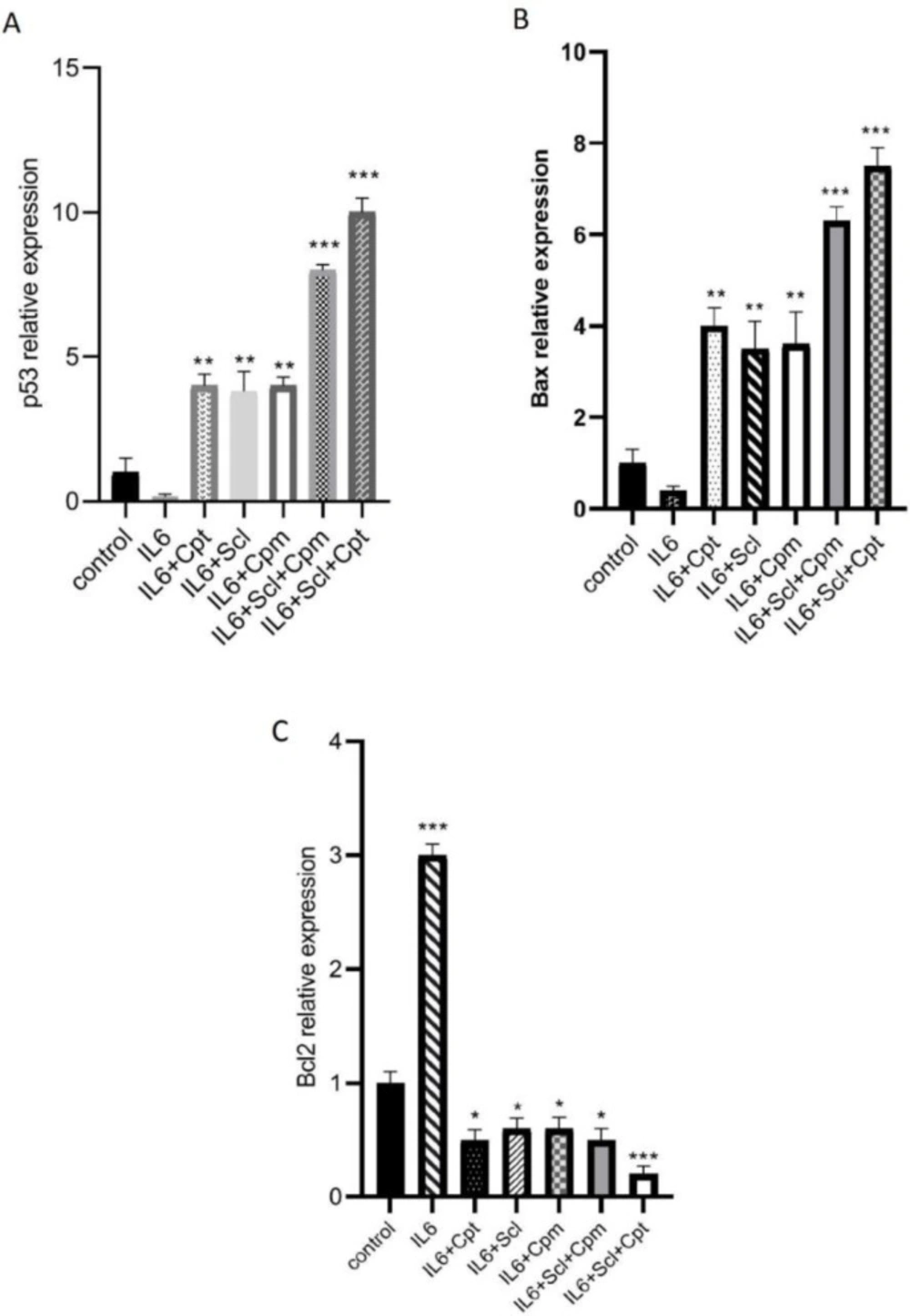
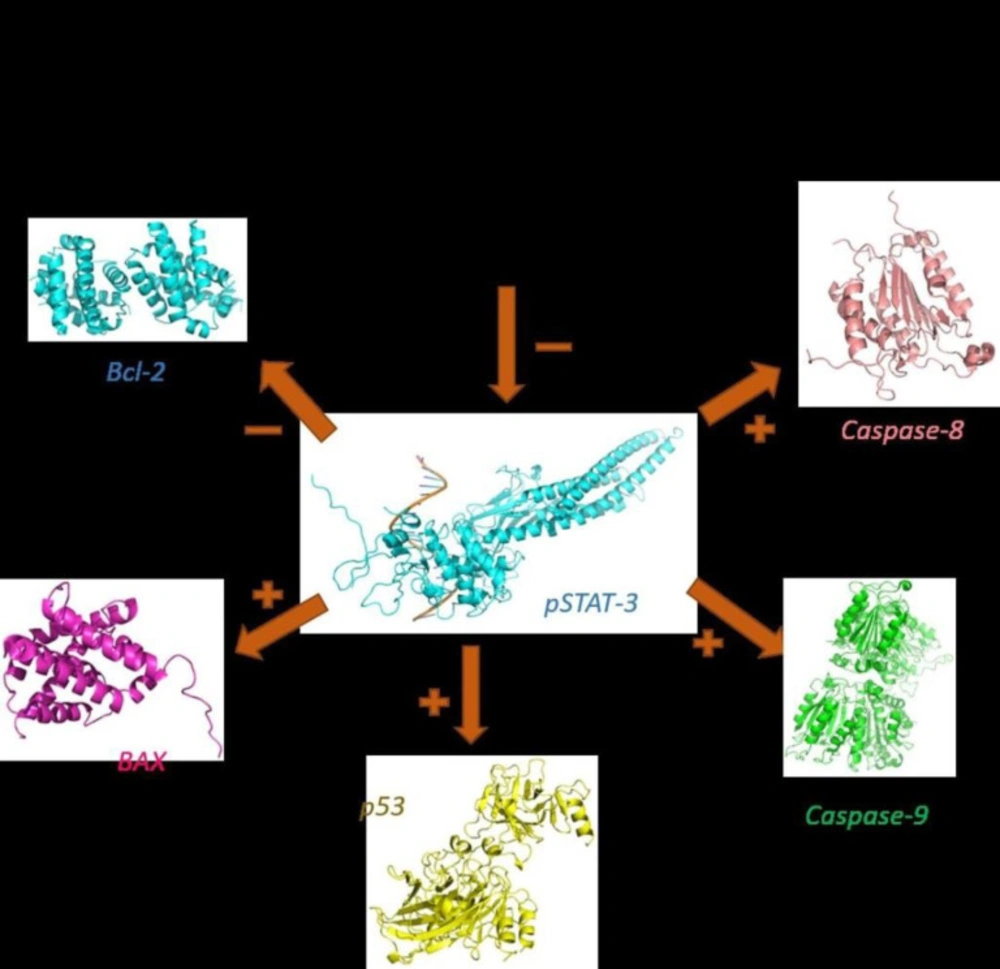
In this study, we showed for the first time that sclareol significantly enhanced the expression of major pro-apoptotic proteins including p53 and BAX and attenuated the expression of Bcl-2 as the chief anti-apoptotic factor (
Figure 8).
In addition to serving as the promoter of metastasis and a regulator of cell invasion (
33,
34), Bcl-2 is known to be involved in the mediation of chemotherapy resistance in some types of cancers (
35-
38). Accordingly, sclareol may have the potential to overcome cancer chemoresistance by inhibiting Bcl-2. Consistently, the up-regulation of BAX and suppression of Bcl-2 by sclareol has been previously reported in osteosarcoma cells (
14). Nevertheless, sclareol has exhibited apoptosis-promoting properties in p53-null cell lines (
29) and so it seems that other pathways might also be involved in the efficacy of sclareol in the induction of cell death. Anyhow, further investigation is required to clarify the detailed mechanism of these effects by sclareol.
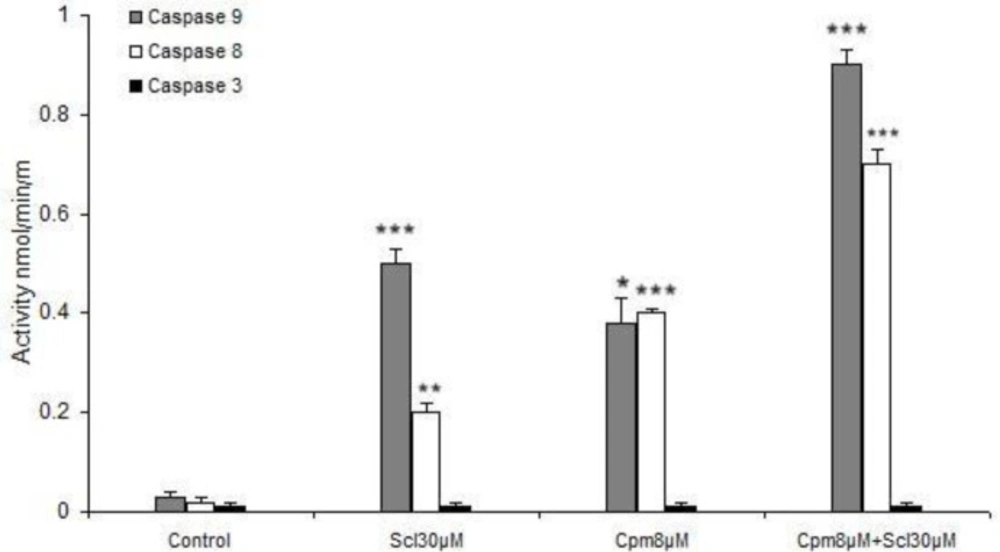
Here we revealed that caspase 8 was activated by sclareol which suggests the stimulation of the extrinsic pathway of apoptosis by this compound. It has been established that in case of high levels of activated caspase 8, the effector caspases 3, 6, and 7 are triggered, while in some other cell types, activation of caspase 8 leads to cytochrome c release from mitochondria which forms the apoptosome complex that results in stimulation of caspase-9 (
39). Caspase-9 has been described as the initiator of the intrinsic pathway that binds to Apaf-1 and cytochrome c and then activates caspase-3 and subsequently leads to apoptosis (
40). We found out that caspases 8 and 9 were activated by sclareol. Thus, based on our findings it seems that activation of caspases in breast cancer cells in response to sclareol follows the latter mechanism and happens through the cross-talk between extrinsic and intrinsic pathways of apoptosis. Caspase 3 is not normally expressed in MCF-7 breast cancer cells (
41), hence its expression was not induced in different experimental conditions.
Meanwhile, the activities of caspase-8 and -9 were also markedly increased in the MCF-7 cell line co-treated with these two compounds comparing to that of either agent alone (
Figure 7). In addition, we observed that the co-treatment more greatly decreased the expression of Bcl-2 and increased expressions of Bax, compared to either agent treated alone.
Sclareol effects on the JAK2/STAT3 path-way
STAT3 is persistently activated in numerous cancers and is involved in cancer cell proliferation, invasion, and migration. Additionally, it tempers epigenetic modification, induces epithelial-mesenchymal transition and promotes cancer stem cells self-renewal and differentiation (
22).
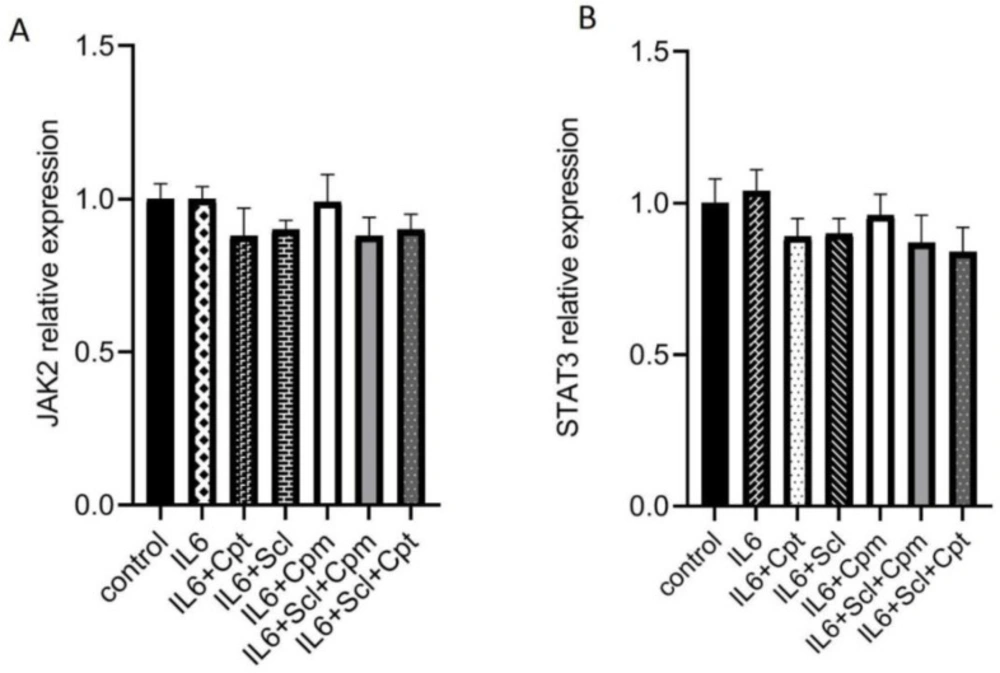
Therefore, the present study aimed to elucidate the molecular mechanism underlying sclareol anticancer activity, particularly in regards to its effects on the JAK2/STAT3 signaling pathway. We showed that sclareol could interact with STAT3 and attenuate its phosphorylation. As reported in previous studies, high concentrations of phosphorylated STAT3 (p-STAT3) was reported to be associated with high metastatic rate of a majority of human cancers (
42-
44) such as lung (
45), ovarian (
46), colorectal cancers (
47) and TNBC (
48) and their poor survival outcomes. In addition, as described previously, accumulating evidence suggests that STAT3, as a critical multifunctional mediator plays a crucial role in the regulation of angiogenesis and cell adhesion under both physiological and pathological conditions (
49-
51). However, the expression levels of non-phosphorylated JAK2 and STAT3 themselves were not affected by sclareol, suggesting that sclareol may intervene principally at the post-translational phase.
Additionally, we found that the modulatory functions of IL-6 on the expression of apoptotic genes were impeded by sclareol. Considering the role of IL-6 on the initiation of the JAK/STAT pathway, inhibition of the IL-6 function by sclareol further verified the effect of this compound on hindering the STAT3 phosphorylation and activity. The addition of cryptotanshinone, a well-characterized inhibitor of STAT3, to the cells treated by sclareol enhanced the inhibitory effect of sclareol and confirmed that the effects of sclareol on breast cancer cells are mediated through JAK/STAT pathway.
Interestingly it was found that combined treatment of sclareol and cyclophosphamide attenuated STAT-3 phosphorylation compared with groups of sclareol and cyclophosphamide treated alone in MCF-7 cell line.
Taken together, sclareol may be considered as a new promising lead for the development of active anticancer agents. However, many of the properties of sclareol are still unknown. Hence, further preclinical and clinical studies are needed to figure out sclareol toxicity as well as its pharmacokinetic profiles, before introducing an officially-marketed medicinal product of this compound.
The effect of sclareol, after 24 h (A) and 48 h (B), cyclophosphamide after 24 h (C) and 48 h (D), or their combination after 24 h (E) and 48 h (F), on the viability of MCF-7 cells. Scl: sclareol; CPM: cyclophosphamide. The data shown are the mean ± SD of at least three separate experiments. The values were compared with the control. Treatment with IL-6 served as positive control
The results of the flow cytometric analysis of the apoptosis in MCF-7 cells treated with sclareol (Scl) (30 µM), cyclophosphamide (Cpm) (8 μM) and their combination, compared with control cells
The effect of sclareol (Scl) alone or in combination with either cyclophosphamide (Cpm) or cryptotanshinone (Cpt) on IL-6-induced modulation of apoptotic genes expression including p53 (A), BAX (B) and Bcl-2 (C). The presented data are mean ± SD of at least three separate experiments. The obtained values were compared with the control. Treatment with IL-6 served as positive control
The effect of sclareol (Scl) in combination with IL-6, cryptotanshinone (Cpt) and cyclophosphamide (Cyc) on the gene expression of JAK2 (A) and STAT3 (B) compared to control group and the cells treated with IL-6. The presented data are the mean ± SD of at least three separate experiments
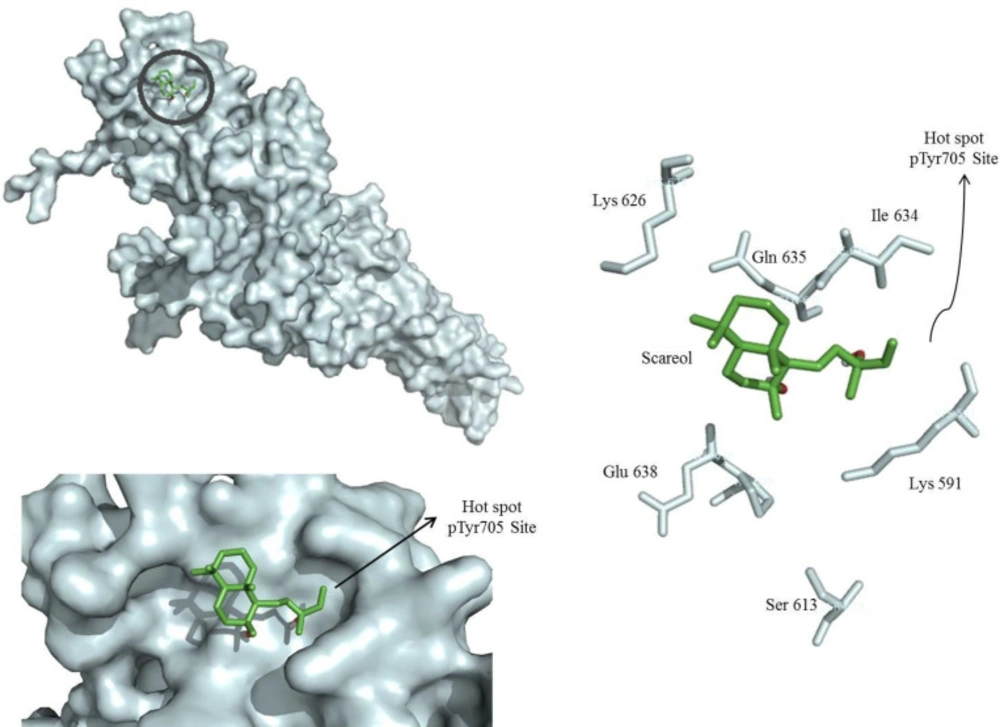
Molecular docking model of sclareol (Scl) binding to the STAT3 SH2 domain (PDB: 1BG1) generated by autodock vina. The surface of the SH2 domain was colored pale cyan and the key residues are shown. Sclareol bound to hot spot pTyr705 site of STAT3 SH2 domain
The phosphorylation status of STAT3 after treatment of cells with sclareol (Scl) either alone or combined with cyclophosphamide (Cyc) or cryptotanshinone (Cpt). Untreated cells that received only the solvent (ethanol) were used as negative control and treatment with IL-6 served as the positive control
Activation of caspases 8 and 9 but not 3 by sclareol (Scl) and cyclophosphamide (Cyc). The presented data are the mean ± SD of at least three separate experiments. * P<0.05, ** P<0.01, *** P<0.001
The pro-apoptotic and anti-apoptotic factors modulated by sclareol in MCF-7 cells
| Primer name | Sequence |
|---|
| JAK2 | F: 5'-GGGAGGTGGTCGCTGTAAAA-3'R: 5'-ACCAGCACTGTAGCACACTC-3' |
| STAT3 | F: 5'-GGTTGGACATGATGCACACTAT-3'R: 5'-AGGGCAGACTCAAGTTTATCAG-3' |
| Bcl-2 | F: 5'-GGTGGGGTCATGTGTGTGG-3'R: 5'-CGGTTCAGGTACTCAGTCATCC-3' |
| BAX | F: 5'-GATGCGTCCACCAAGAAGC-3'R: 5'-AAGTCCAATGTCCAGCCCAT-3' |
| p53 | F: 5'-CAGCACATGACGGAGGTTGT-3'R: 5'- TCATCCAAATACTCCACACGC-3' |