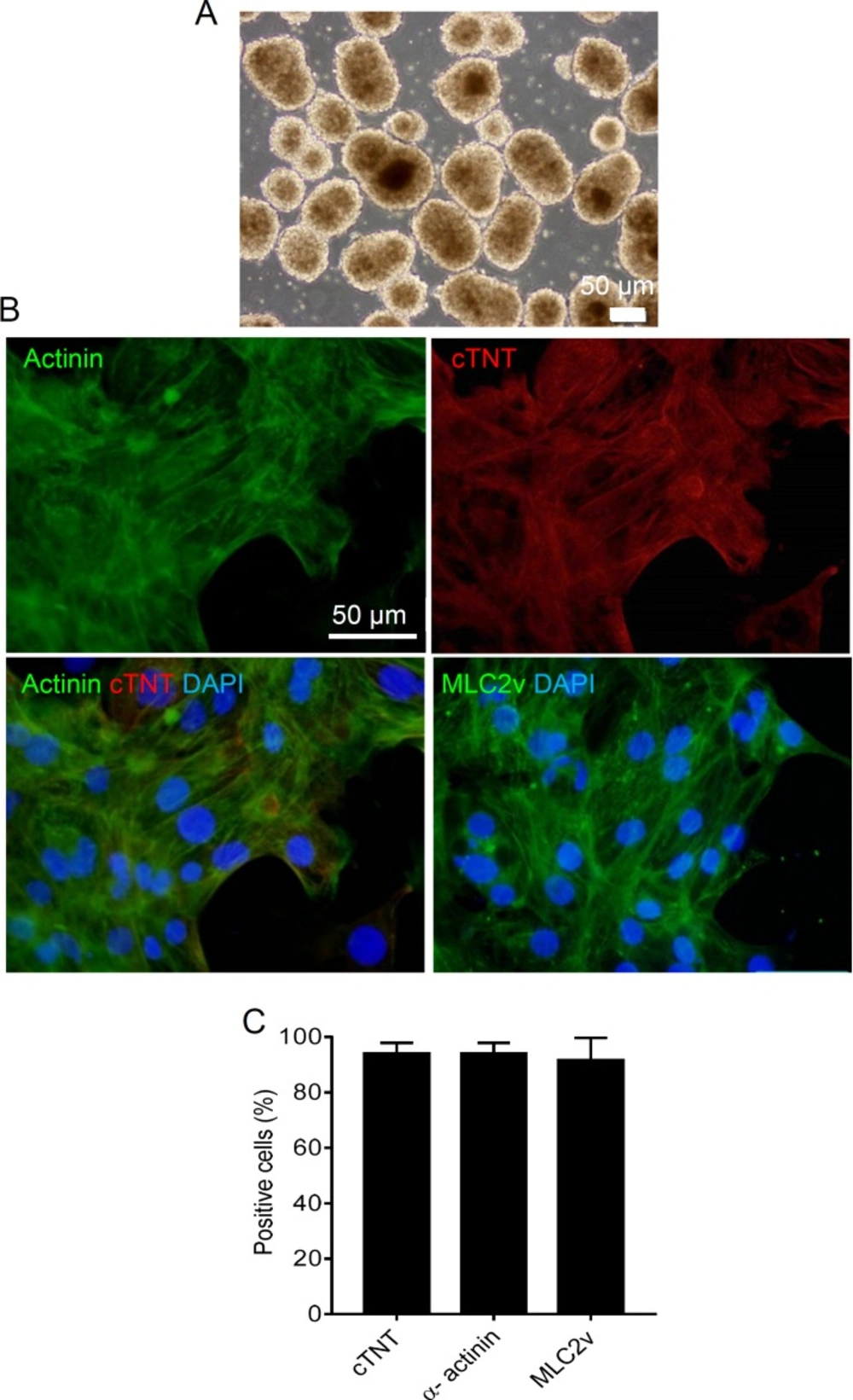
CDCs have shown to promote cardiac regeneration of the infracted human heart (
2,
17). Exosomes generated by CDCs are beneficial paracrine signals that reproduce CDC-induced therapeutic regeneration. They are sufficient to mediate the entire effect of CDCs (
3,
5). Few studies have investigated the therapeutic potential of CDC-exosomes in animal MI models and some other cardiovascular diseases. These studies have shown that exosomes secreted by CDCs replicate the cardioprotective and regenerative effects of CDCs such as apoptosis inhibition of cardiomyocytes. Gallet and coworkers indicated that CDC-derived exosomes delivered by intramyocardial (IM) injection has the ability to decrease acute ischaemia-reperfusion injury, halt adverse remodeling and to improve LVEF in pig models of acute (AMI) and convalescent myocardial infarction (CMI) (
2). In the study of Ibrahim
et al., exosomes secreted by human CDCs inhibit apoptosis and promote proliferation of cardiomyocytes, while enhancing angiogenesis. Injection of exosomes into injured mouse hearts recapitulates the regenerative and functional effects produced by CDC transplantation, whereas inhibition of exosome production by CDCs blocks those benefits (
3). All of these data confirmed
in-vivo anti-apoptotic effect of CDC-exosomes (
2,
3,
18).
In all of these studies, CDC-exosomes were obtained under normoxic conditions, which likely could not reflect the state of post-infarct tissue. While most
in-vitro cultured cells maintained at oxygen levels of approximately 20%, natural cell micro-environments seem to have much lower oxygen tensions with considerable variation based on location. For instance the mean oxygen concentration of arterial blood is approximately 12%, and that of tissue is 3% (
19). Adult stem cells similarly live under hypoxic conditions of 3-5% O
2in-vivo and these hypoxic conditions are the physiological norms for a variety of stem cell niches (
20). These studies have shown that the level of oxygen play a crucial role in the maintenance, differentiation, and function of stem cells. Nevertheless, hypoxia can also induce mitochondria-mediated apoptosis and subsequent caspases activation in bone marrow-derived mesenchymal stem cells (
21). In this study we have analyzed the anti-apoptotic activity of exosomes generated by CDCs under both normoxic and hypoxic conditions. Here, we showed that H-exo and N-exo significantly decrease CoCl
2-induced apoptosis in hESC-CM.
Here, to isolate the CDC-exosomes from conditioned media, differential ultracentrifugation was used as described in the literatures with some modifications (
12,
13). The size of H-exo and N-exo were almost similar, with mean hydrodynamic diameter of 150 to 170 nm. CDC-derived exosomes possessed highly positive expression for exosome surface markers, such as CD63 and CD81 (unpublished data).
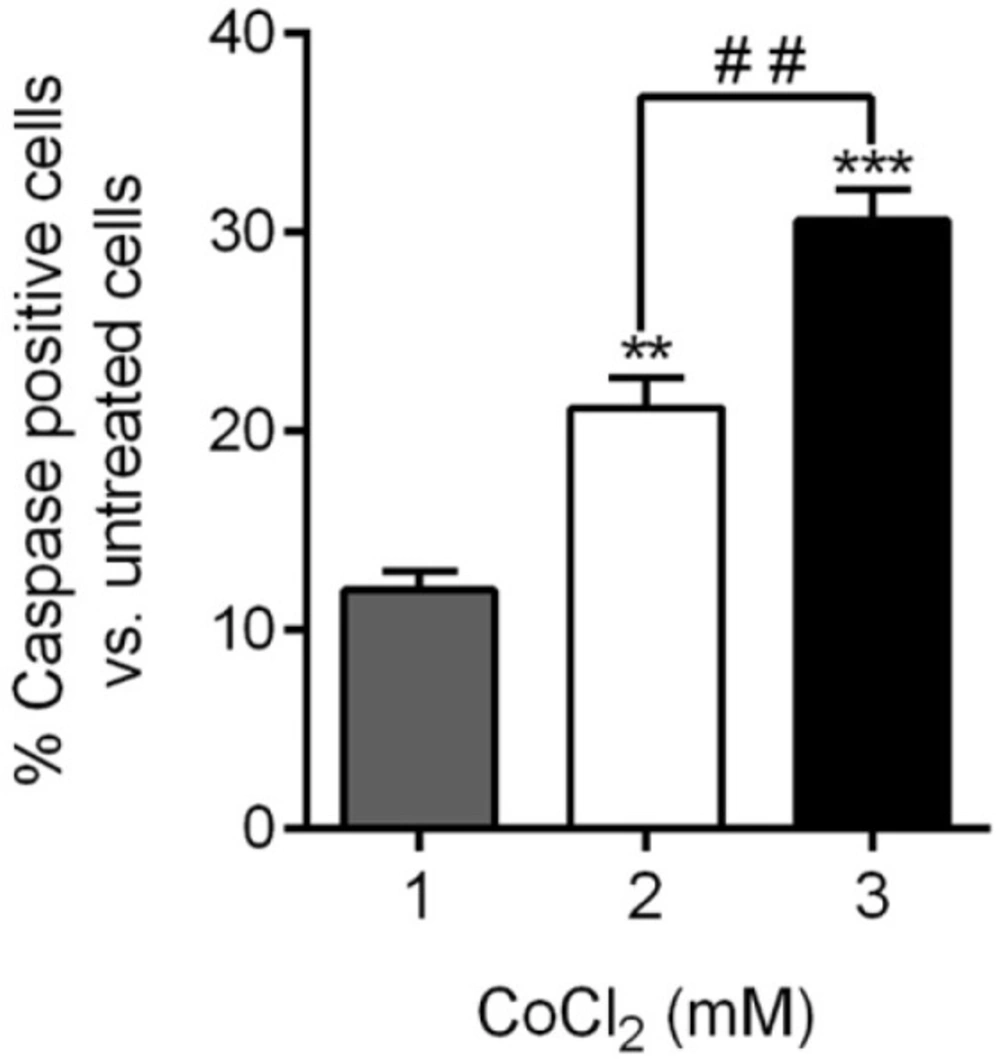
CoCl
2 is a well-established hypoxia-mimicking substance. CoCl
2-treated cells share common features with cells incubated at 1% oxygen (
22). CoCl
2, as a substrate of the ferrochelatase enzyme, is thought to mimic the hypoxia by binding to the heme molecules (instead of Fe
2+). It was shown that the expression level of hypoxia-inducible factor-1α (HIF-1α), which is a major transcription factor and key regulator of adaptive responses to hypoxia, is markedly increased following treatment with CoCl
2 in a dose-dependent manner (
23). In this study, CoCl
2 at concentration of 3 mM was used to induce apoptosis. In the study of Guo
et al., U937 and NB4 the cell lines were treated by CoCl
2 at different concentrations of 150, 200 and 300 μM. They found that at the concentration of 150, 200, and 300 μM the viability is reduced to 55, 20, and 7 % in U937 and to 60, 50, and 25% in NB4, respectively (
15). Kim
et al. also demonstrated that neural cells viability is reached to about 60% after 24 h CoCl
2 treatment at concentration of 1 mM (
16). However, it should be taken into consideration that the method of apoptosis detection is not the same in all studies. For example, trypan-blue exclusion and MTT assay were used to evaluate cell viability by Guo (
15) and Kim (
16)
et al., respectively. Totally, our data are in agreement with the results of Guo
et al. (
15) which indicate CoCl
2 at the concentration of greater than 50 μM induce apoptosis via mitochondria pathway-mediated caspase-3 activation. (
15).
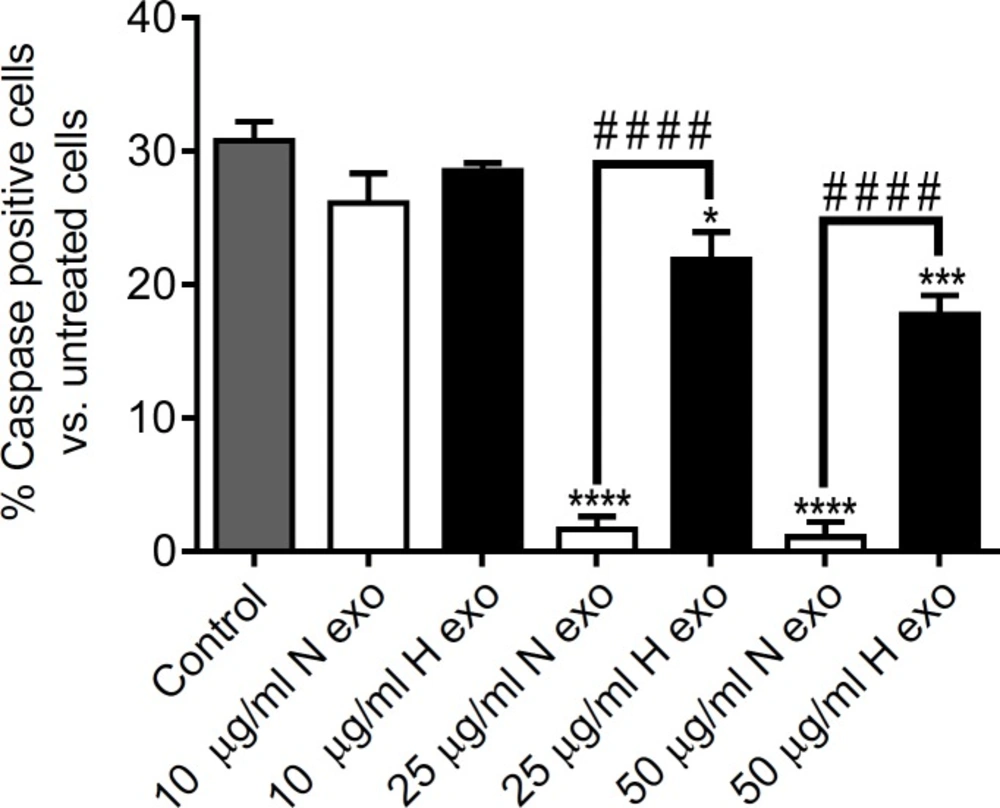
In this study, we found that exosomes secreted by human CDCs were cultured 48 h under hypoxic and normoxic conditions (1% O
2) inhibited apoptosis at both 25 and 50 μg/mL concentrations. However, the anti-apoptotic effect of N-exo was significantly higher than that of H-exo at concentration of 25 and 50 μg/mL (
p <0.0001). In the present study, higher oxygen percentage (1% O
2) was used for hypoxic preconditioning compared to hypoxic culture condition (0.5% O
2) that was used in the study of Chacko
et al. (
24). Their results indicated that exposure to sub-lethal hypoxia (0.5% O
2) for as long as 72 h by itself does not induce cell death by apoptosis in mesenchymal stem cells (MSCs) (
24). Furthermore, exposure of hypoxic preconditioned MSCs (HPC-MSCs) (0.5% O
2, 24 h) to severe hypoxia (0.1% O
2, 48 h) resulted in significant decrease in apoptosis compared with non-HPC MSCs (
24). In another study, to imitate myocardial ischemic–reperfusion, isolated MSC were induced by hypoxia/reoxygenation (H/R) [6 h hypoxia (<0.5% O
2) followed by 12 h reoxygenation (21% O
2)]. It was found that hypoxia preconditioning (8% O
2 hypoxia and 30 min of reoxygenation) inhibits H/R-induced apoptosis of MSC in a time-dependent manner (
25). Khatibi
et al. reported that preconditioning with H
2O
2 along with CoCl
2 can cause inhibitory effect on cell apoptosis (
26).
Recently, a study has reported that the CPC-derived exosomes might be crucial to protect the cardiomyocytes from apoptosis caused by oxidative stress. In their study, exosomes secreted by mice CPCs which were stressed by 100 μM H
2O
2 for 6h (H
2O
2-exosomes) decreased the percentage of the apoptotic cells to 13.58%, compared with the 33.29% in H
2O
2 group, (cardiomyocytes were stressed by 100 μM H
2O
2 for 6 h), whereas the normal exosomes (non-H
2O
2 induced) could only reduce the apoptotic percentage to 17.39% (
27).
Totally, it can be concluded that different parameters in hypoxia preconditioning of cardiac stem cells including the percentage of oxygen and the duration of hypoxia play critical roles in their anti-apoptotic effect. Therefore, to obtain the optimum anti-apoptotic effect of cardiac stem cells (CSCs)-derived exosomes, further investigation is highly required to choose the proper manner of hypoxic preconditioning.
Additionally, the way of apoptosis induction in cardiomyocyte might be important in the anti-apoptotic effect of CSCs-derived exosomes. The severe hypoxic (0.1% O
2) and reduced serum conditions that led to decreasing cells apoptosis in the study of Chacko
et al., (
24) may be different from the apoptosis condition that was induced by cobalt chloride (3 mM) in our study
. In the study of Xiao
et al., H
2O
2 was used to induce the oxidative stress that originates mainly in mitochondria from reactive oxygen species (ROS) (
27,
28). Their results demonstrated that H
2O
2-exosomes reduce H
2O
2 induced apoptosis. Further investigations dealing with the kind of apoptosis induction will be helpful.
We can get to the conclusion that CDC-secreted exosomes have the potential to prevent apoptosis in cardiomyocytes and they will hopefully provide a promising therapeutic strategy for ischemic cardiac disease. Our results imply the need for further investigation of the effect of hypoxia-preconditioning method of cardiac stem cells on the anti-apoptotic activity of their secreting exosomes.