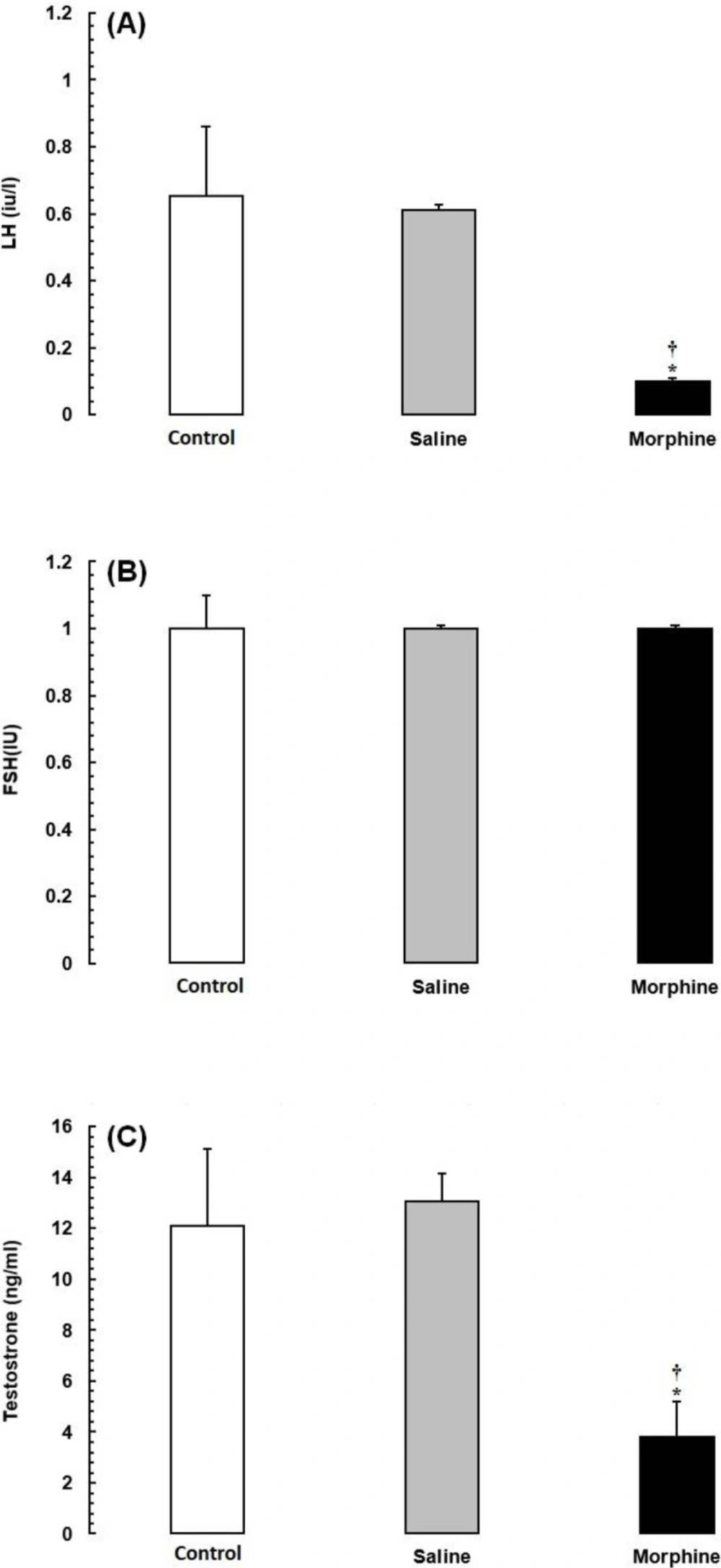
Our study detected significant associations of opiate addiction with the impairment of male fertility-related parameters and decreased markers in fertility-related pathways. Besides, the serum LH and testosterone levels were significantly lower in the addicted rats than in the saline and control rats. Despite the presence of hypotestosteronemia, low LH, and impaired spermatogenesis, the serum level of FSH did not increase in the addicted rats. These results are consistent with previous studies demonstrating that opiate consumption can result in a state of hypogonadotropic hypogonadism (
26). In another similar study, it was shown that the administration of morphine to male rats led to a decrease in serum levels of LH and testosterone and decreases in the testis weight and sperm production (
27). A subsequent study by Lee
et al. in 1978 on addicts confirmed the lowering of the blood testosterone levels, but changes in the FSH and LH levels were not determined at that time (
28). In animal studies conducted on rats and pigs, there was no change in the FSH levels despite a significant decrease in the LH and testosterone levels (
29). Drug use appears to change the sexual function and the level of associated hormones (
29). A decline in male fertility has occurred in recent years, with one of the main reasons for increased exposure to toxicants in the environment. These agents may be chemical materials, stress, and ionizing radiation, as well as substance abuse (
30,
31). In line with our data, this study showed that administration of morphine also reduced serum reproductive hormones and confirmed that the damage induced by morphine caused to spermatogenesis.
Since previous studies have investigated the effect of opioids on male infertility, they examined various molecular pathways in testicular tissues. They showed that drug use was effective on pathways involved in infertility. In our present study, the morphine group had a significantly lower expression of male fertility genes. In addition, this finding implies that the impaired male fertility found in some cases could be due to drug influences in the hypothalamus and pituitary gland anda direct effect on the DNA integrity in testis tissue. The genes and proteins related to spermatogenesis and metabolism can be effective in normal sperm function. One of these genes is
HDAC1, which is present in the nucleus of the Sertoli cells responsible for producing sperm. On the other hand, opiates have receptors in different parts of the testicles, including Sertoli cells and seminiferous tubules, which confirms their direct impact on spermatogenesis. They affect the
HDAC1 gene in the nucleus of Sertoli cells and disrupt the spermatogenesis cycle; this gene has a critical role in regulating the changes in the major histones and in the control of chromatin changes that affect the transcription of tumor suppressor genes, causing tumorous and pathological changes in the tissue (
20).
HDAC1 is linked to different pathways, including the mTOR, MAPK, Notch, Hippo and Wnt signaling pathways associated with cell growth, proliferation and differentiation (
32).
Different studies have shown that disruption in each of these pathways is associated with infertility in both men and women (
33,
34). A study conducted by Jee Hyun Kim in 2014 showed that the level of
HDAC1 expression in men with azoospermia was lower than that of normal people and attributed it to the reduction of
HDAC1 gene expression with sperm DFI (
35).
The assessment of
CREB, FOS in our study showed that its protein expression was diminished in the addict animal which is in agreement with a previous research Elena H. Chartoff
et al. suggested that decrease
CREB1, FOS activation in portions of the striatum is related to addict by morphine (
36). The gene transcript for CBP that is a protein binds to
CREB (CBP or Crebbp) and strongly linked protein p300 are essential cofactors for many nuclear transcription factors (
37). Since direct pathways for CBP activation by GnRH and insulin,
i.e., mitogen-activated protein kinase (MAPK) and PKC can phosphorylate CBP, CBP action in the gonadotroph was studied (
38). As FSH increases, it affects the Sertoli receptors and increases the adenylate cyclase and cAMP levels, resulting in phosphorylation of the
CREB1 transcription factor on serine-133 (
39).
CREB1 induces the transcription factors required to activate other genes involved in spermatogenesis (
40). It regulates the hormones that regulate spermatogenesis and anti-apoptotic factors (
41). According to the data shown by the String database, there is an interaction between
FOS with Interleukins receptor, apoptosis-related cysteine peptidase (CASP1), Mitogen-activated protein kinase 14 (MAPK14), Jun proto-oncogene (JUN), and FBJ murine (
42); in addition, according to another study, in evaluating specific types of male infertility, Interleukins and
FOS in seminal plasma should be extended; Thus, the changed
FOS influences the innate immunity level in male infertility (
43).
Studies have shown that
CREB1, CREM, and
FOS in the mouse and rat spermatogonial stem cell model helps the process of spermatogenesis by activating the Ras/Erk1/2 pathway and CDK2 promoter (
44,
45). For expressing the late spermatogenic genes, there is a known master switch, including several spermatid-specific transcriptional regulators. The transcriptional activator CREM
T, highly expressed in round spermatids is essential for expressing many important postmeiotic genes, for instance,
Prm1,
Prm2,
Tnp1 and
Tnp2, is encoded by the
Crem gene (
46,
47).
The coordinated action of a set of general transcription factors is necessary for transcription initiation by RNA polymerase II in eukaryotes (
48,
49). The
TBP is one of the major factors in transcription initiation; as well, during spermatogenesis, it shows a stage-specific expression pattern (
50). Previous studies showed that testicular tissue in sterilized male mice was investigated, and the number of genes involved in spermatogenesis. The number of sperms in these mice was also evaluated. This study showed that spermatozoa were decreased and spermatogenesis was controlled by the
TBP gene and
TBP has been shown to affect the various factors involved in spermatogenesis (
51).
Vesselin M. Chorbov et al. (2011) showed that DNA methylation in addicted men’s sperm was higher than those who quit the addiction. Hypermethylation of CpG loci in the promoter of genes involved in suppressing necrotic tumors, such as HDAC1 and HDAC1 gene expression, decreases. Increased DNA methylation in sperm may represent a method of epigenetic inheritance of opioid abuse or dependence phenotypes (
52). According to Betina González in 2018, drug addiction modifies epigenetic homeostasis and next-generation outcomes, increases methylation of cytosine levels in sperm DNA and germ cells, and decreases HDAC1 gene expression. It also decreases spermatogenesis, which has a higher gene expression rate with drug withdrawal but is lower than normal (
53). Jinghua Wang (2007) stated that morphine chronically inhibits interleukin-2 (IL-2) at the genomic and protein levelresulting in the inhibition of the CREB gene. In addition, chronic morphine treatment inhibits acetylation and trimethylation of histones and decreases DNA demethylation and access to the IL-2 promoter. These findings suggest that chronic morphine treatment may act through both transcriptional and epigenetic mechanisms to inhibit the IL-2 production and ultimately reduce CREB gene expression (
13). Alcohol, cocaine, and nicotine increase methylation of cytosine levels in DNA and decrease FOS gene expression (
54). According to another study, cellular and molecular examination in mice showed that spermatogonia stem cells were necessary for sperm production. Also, knocking down a set of genes, especially the
TBP gene, reduces the production of sperm, which also increases apoptosis rates in these rats (
55).
Effects of morphine on serum reproductive hormone levels in Wistar rats. Data are expressed as the mean ± SEM; *Significant decrease of the hormone level in the morphine group compared with the saline and control groups (P < 0.05).
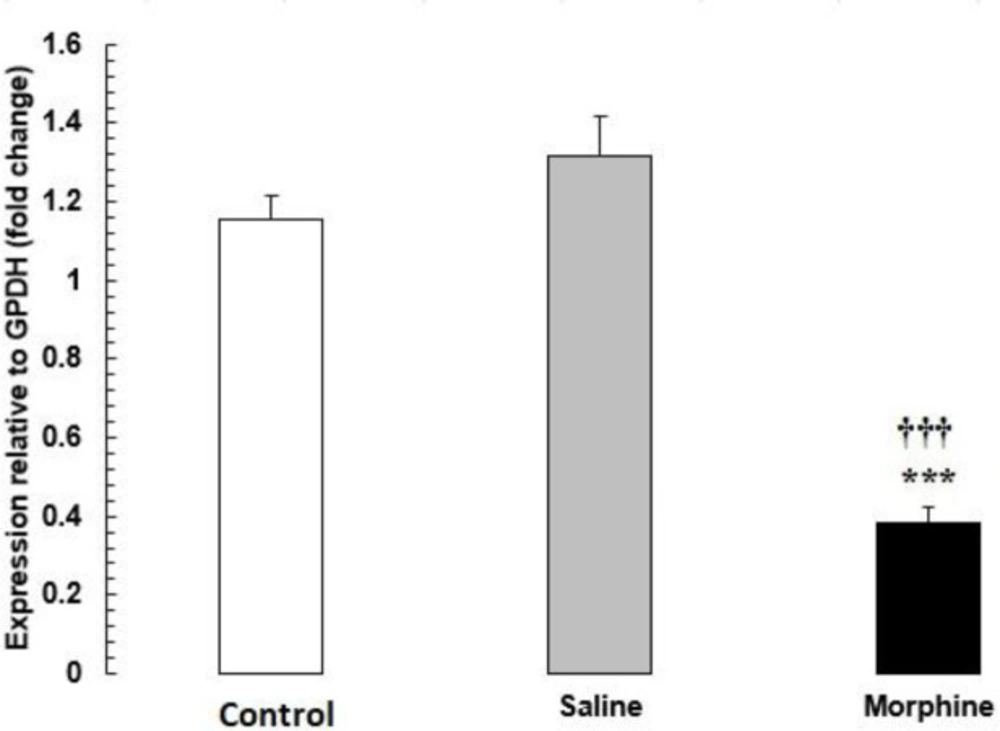
HDAC1 mRNA expression in testis tissue from male rats addicted to morphine. The mRNA amounts were evaluated by a quantitative real-time reverse-transcription polymerase chain reaction. Data are the mean ± SEM (n = 10 for each group). Glyceraldehyde-3-phosphate dehydrogenase was used as an internal control. ***P < 0.001, morphine group vs. saline and control groups
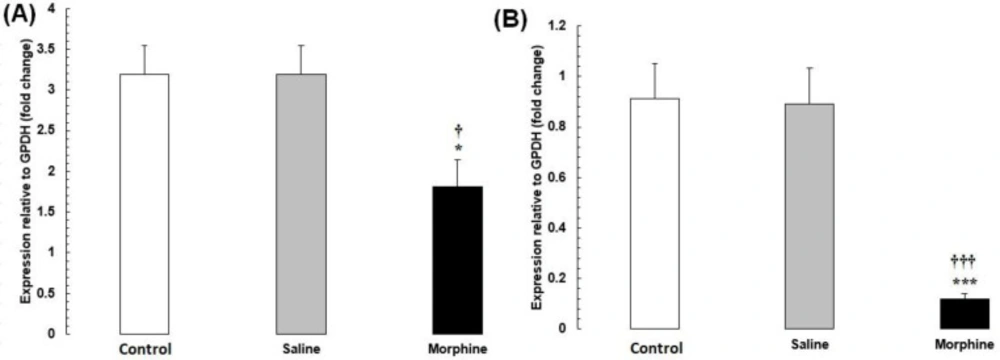
(A) CREM and (B) TBP mRNA expression in testis tissues from male rats addicted to morphine. The mRNA amounts were evaluated by a quantitative real-time reverse-transcription polymerase chain reaction. Data are the mean ± SEM (n = 10 for each group). Glyceraldehyde-3-phosphate dehydrogenase was used as an internal control. *P < 0.05 and ***P < 0.001, morphine group vs. saline and control groups
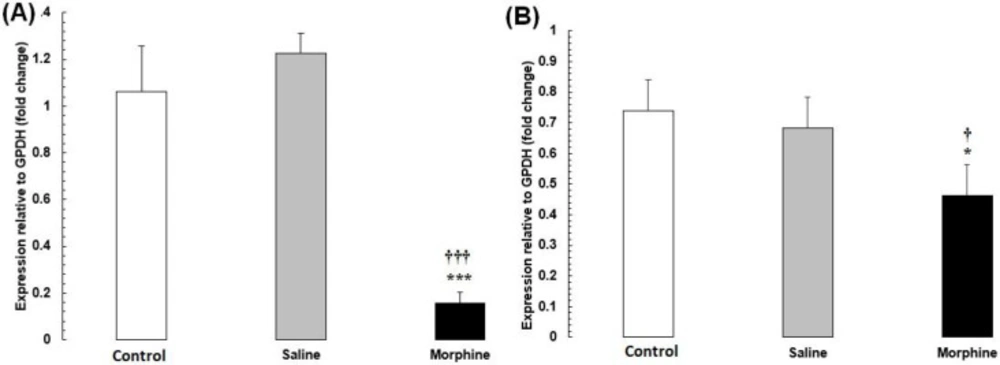
(A) CREB1 and (B) FOS mRNA expression in testis tissues from male rats addicted to morphine. The mRNA amounts were evaluated by a quantitative real-time reverse-transcription polymerase chain reaction. Data are the mean ± SEM (n = 10 for each group). Glyceraldehyde-3-phosphate dehydrogenase was used as an internal control. *P < 0.05 and ***P < 0.001, morphine group vs. saline and control groups
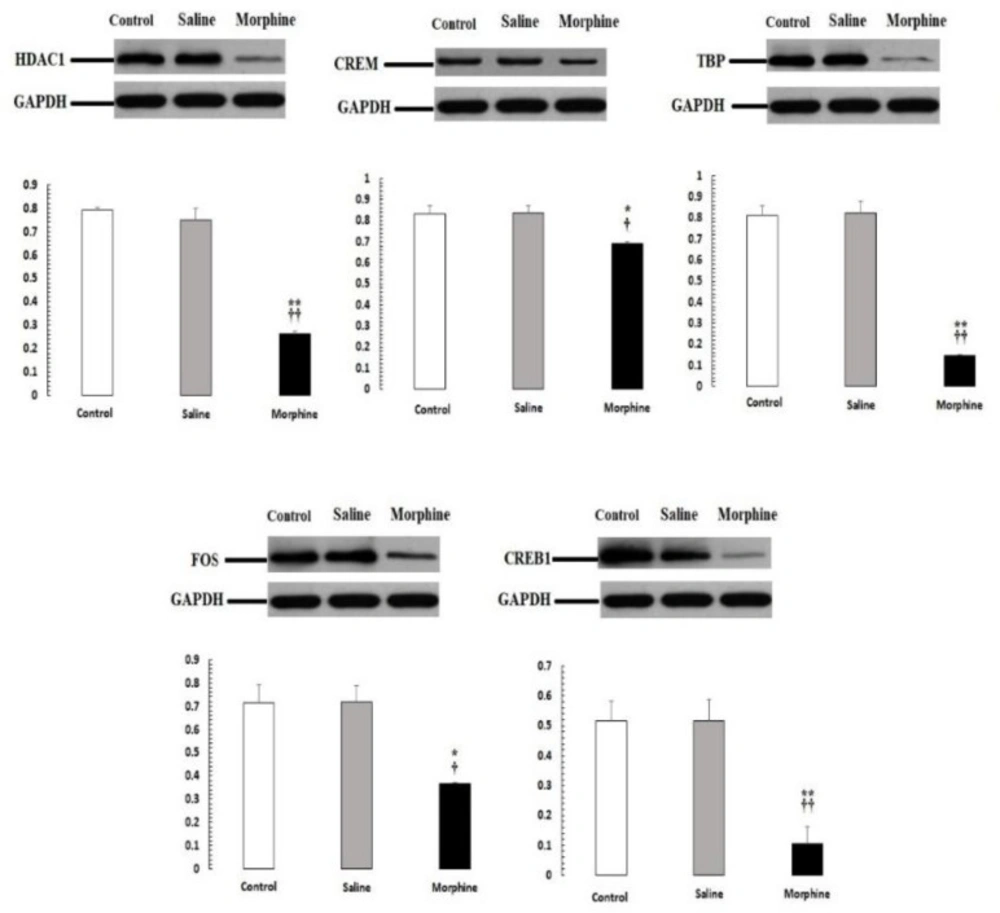
CREM, TBP, CREB1, HDAC1, and FOS protein levels were evaluated by Western blot. Data are the mean ± SEM (n = 10 for each group). Glyceraldehyde-3-phosphate dehydrogenase was used as an internal control. *P < 0.05 and **P < 0.01, morphine group vs. saline and control groups
| Gene | | Sequence (5'->3') | Length | Tm | GC (%) | Self-complementarity | Self-3' complementarity |
|---|
| HDAC1 | Forward primer | GACGGGGATGATGGAAACTAC | 21 | 58.16 | 52.38 | 3.00 | 0.00 |
| Reverse primer | GTTGGATTTGTGAGGACGATAG | 22 | 56.87 | 45.45 | 2.00 | 2.00 |
| CREM | Forward primer | GAAACAACATAGGGTAGAAAGGG | 23 | 56.85 | 43.48 | 2.00 | 0.00 |
| Reverse primer | GAAAATGAGCACAACACTGGATG | 23 | 58.77 | 43.48 | 4.00 | 1.00 |
| TBP | Forward primer | ATCTTCATCCTTGTCCTCCAGCTTC | 25 | 62.38 | 48.00 | 4.00 | 0.00 |
| Reverse primer | GCTCCCTCCAAAGCAATCTTCCTTA | 25 | 62.85 | 48.00 | 6.00 | 4.00 |
| FOS | Forward primer | GGTCCTGTCTGGTTCCTTCTATG | 23 | 60.12 | 52.17 | 3.00 | 0.00 |
| Reverse primer | CTGCCTTGTCTGACTGCTCAC | 21 | 61.21 | 57.14 | 5.00 | 1.00 |
| CREB1 | Forward primer | CAG TTG TTA TGG CGTCCT | 18 | 54.61 | 50.00 | 2.00 | 2.00 |
| Reverse primer | CTT GCT GCT TCC CTG TTC | 18 | 55.99 | 55.56 | 3.00 | 0.00 |
| GAPDH | Forward primer | CAT ACT CAG CAC CAG CAT CAC C | 22 | 61.32 | 54.55 | 3.00 | 0.00 |
| Reverse primer | AAG TTC AAC GGC ACA GTC AAG G | 22 | 61.58 | 50.00 | 5.00 | 0.00 |