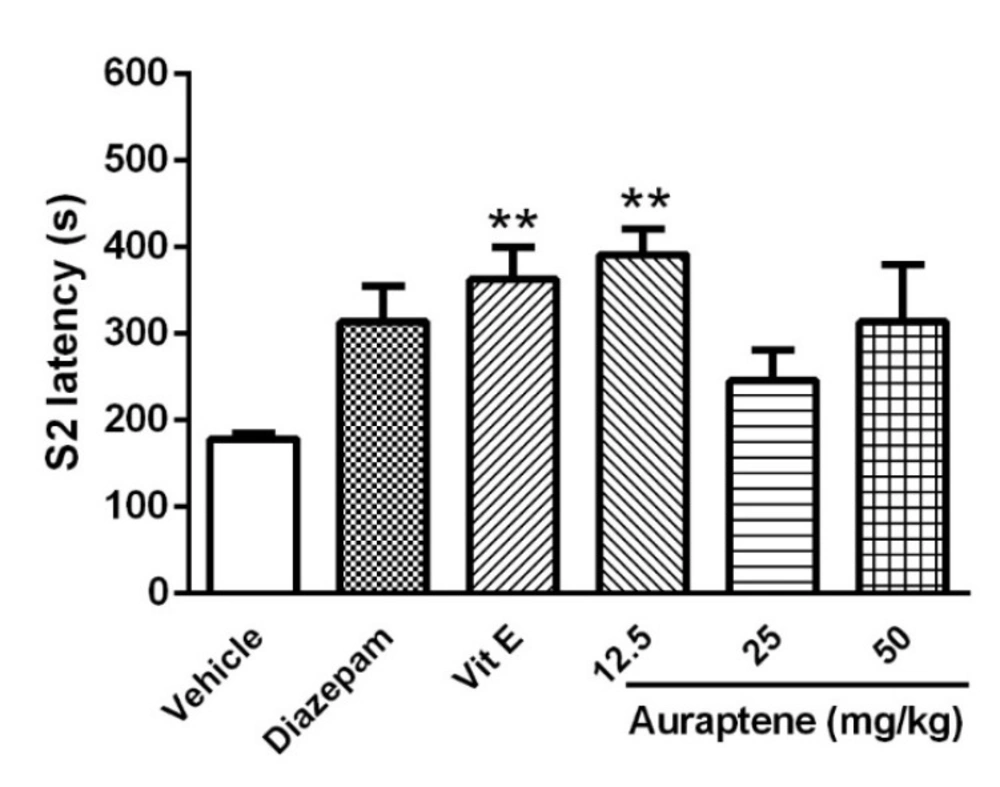
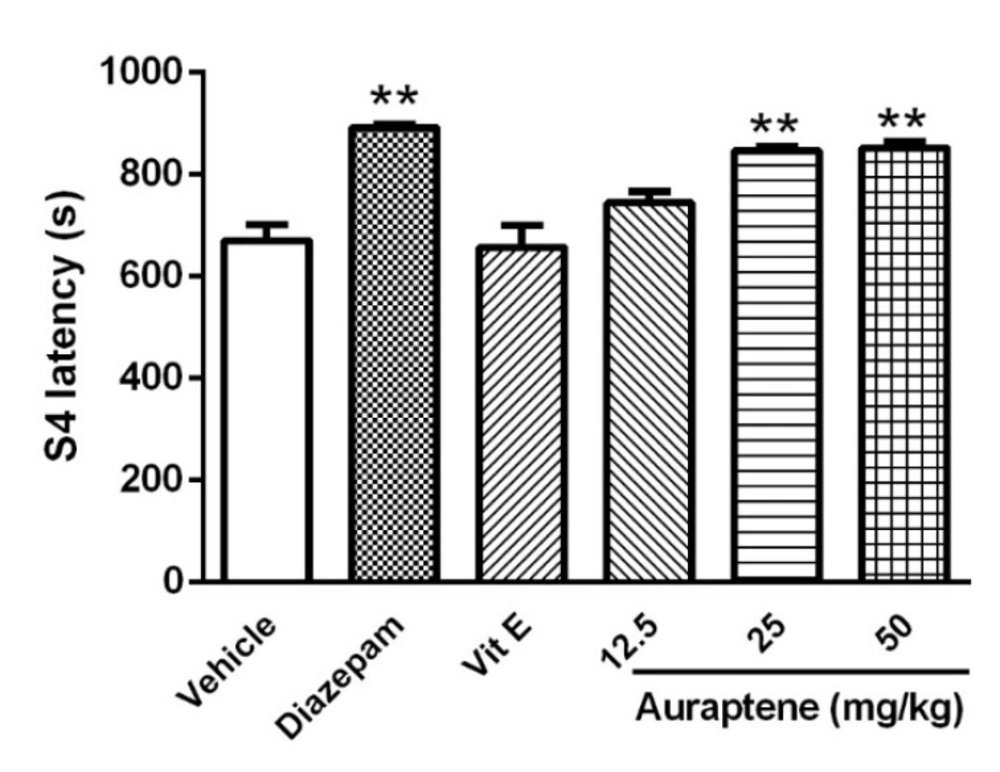
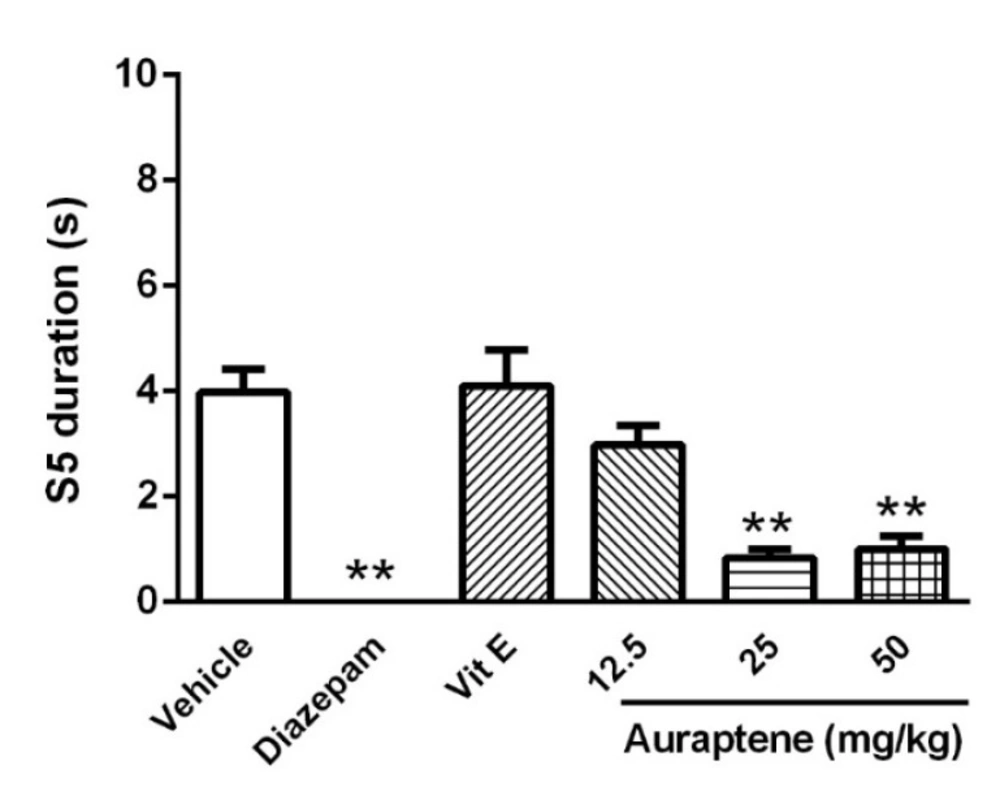
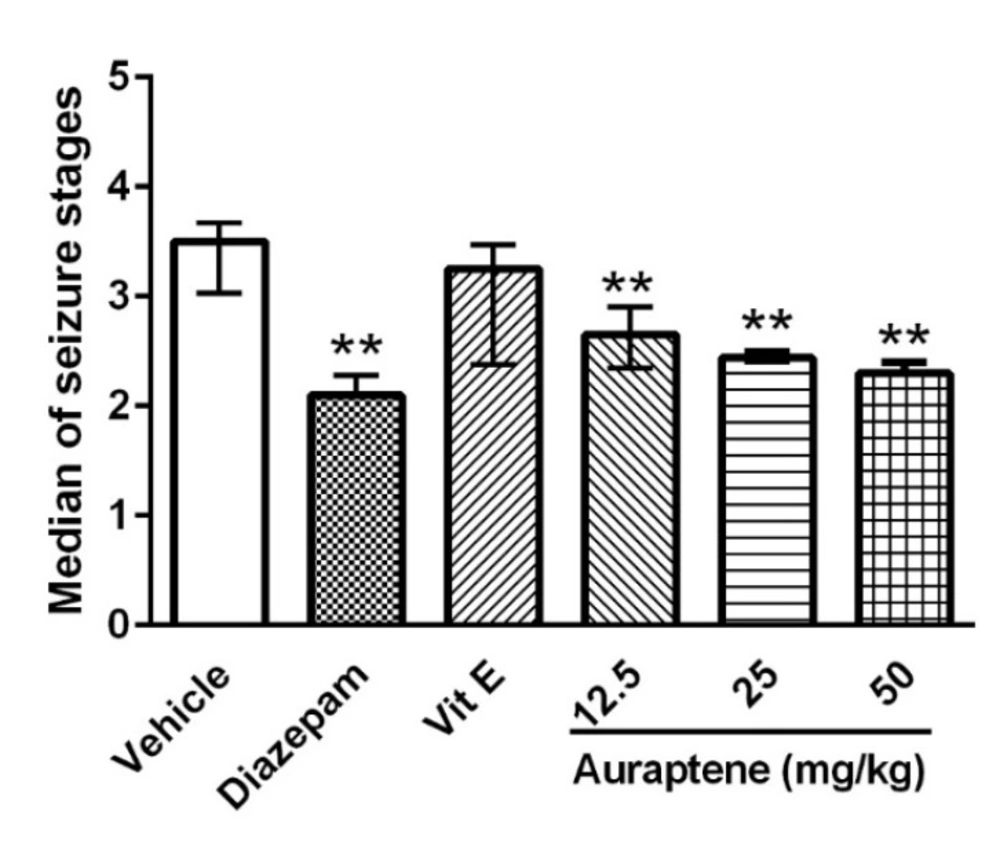
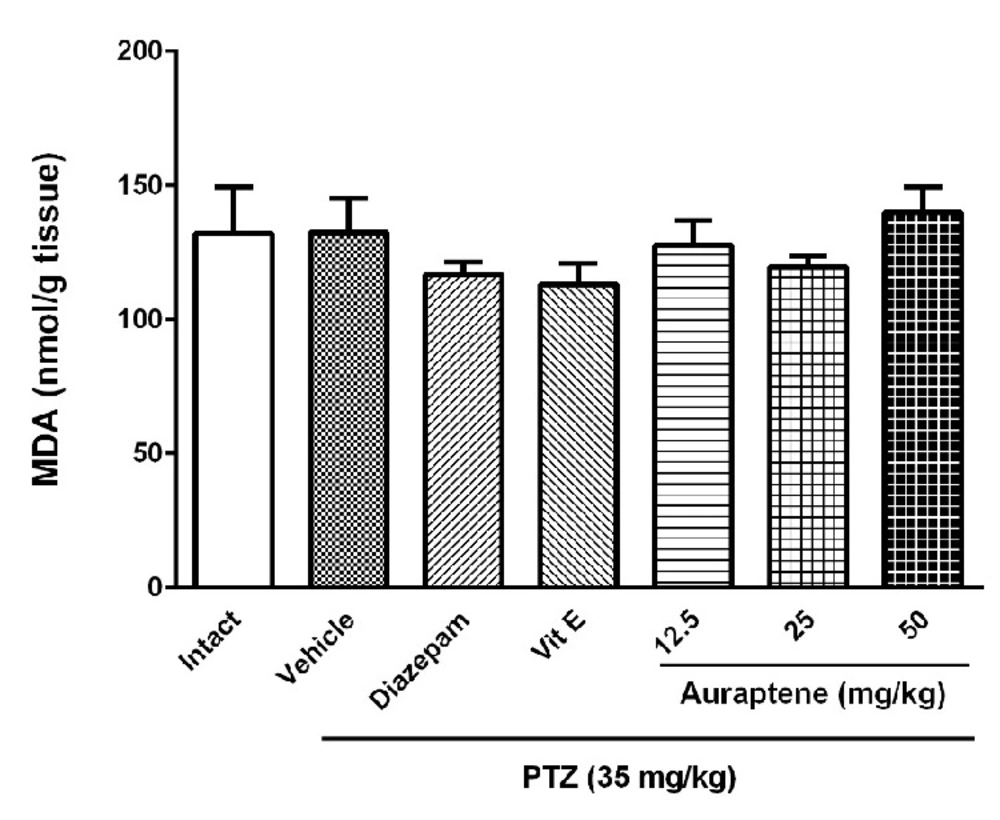
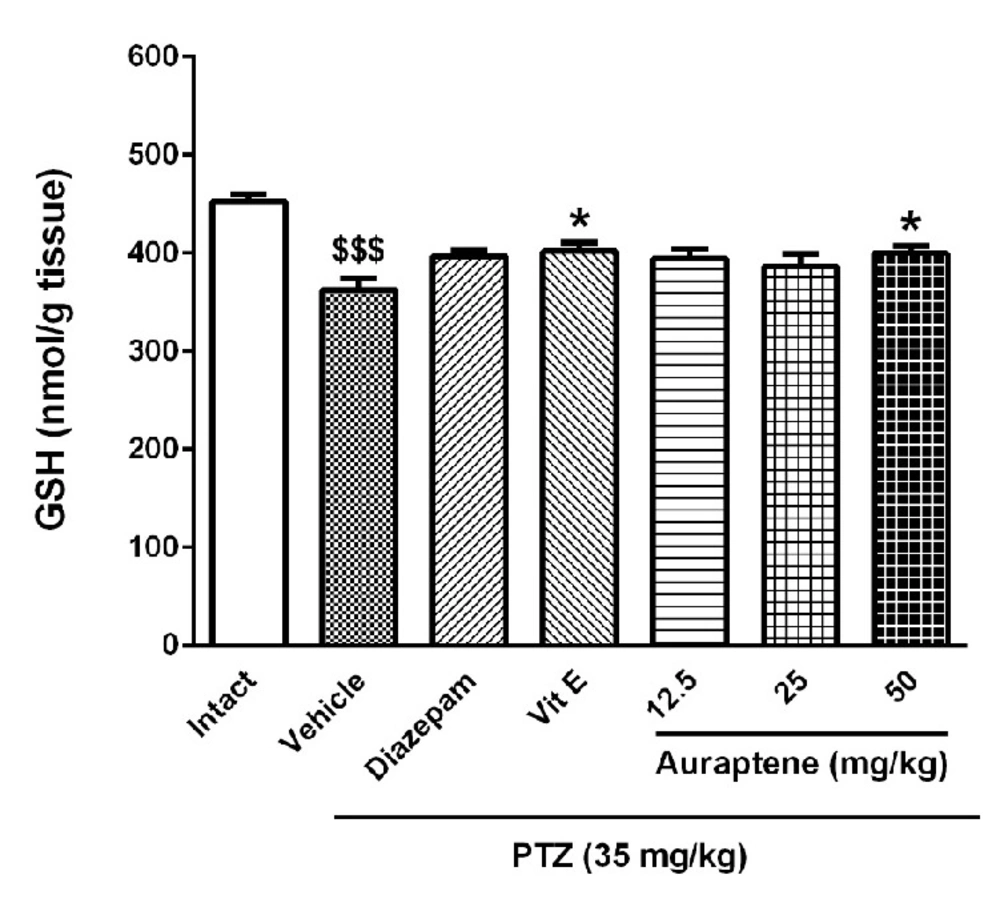
The main finding of the present study is that auraptene had anticonvulsant effect in PTZ kindled mice. It also increased reduced glutathione in the brain similar to vitamin E. However, vitamin E did not produce such anticonvulsant effects in PTZ kindled mice. Compelling evidence from previous studies has shown that some natural compounds have significant anticonvulsant effects (
17-
19). Coumarins are polyphenolic compounds that possess a diverse range of pharmacological effects. It was reported that administration of PTZ increased free fatty acids, glutathione disulfide, and hydroxyl radicals in various brain regions including the cerebral cortex (
8,
20). Also, epileptic patients have been reported to have lower plasma vitamin C content and higher levels of lipid peroxidation in comparison with normal subjects (
21). Auraptene, as an herbal compound, was able to decrease main antioxidant enzymes activities including the superoxide dismutase and glutathione peroxidase (
9). Therefore, we hypothesized that oxidative stress might have a potential role in PTZ-induced chemical kindling and auraptene would be able to resolve these biochemical changes. The results showed that induction of kindling reduced GSH content in the brain but did not change the MDA level significantly. Similar to these results, Patsoukis
et al. showed that a single injection of PTZ reduced GSH level in the cerebral cortex of mice but did not affect the MDA levels (
8). The present finding is in accordance with previous studies showing that a perturbation in oxidant-antioxidant balance is accounted, at least in part, as a player in the pathogenesis of seizure. Although vitamin E enhanced GSH and decreased stage 2 of seizure attacks, it did not change the median of seizure scores and higher stages of the seizure. Therefore, vitamin E, as a good antioxidant molecule, did not induce a prominent anti-epileptic effect. In other words, vitamin E exhibited a weak anticonvulsant effect. In agreement, a recent study showed that N-acetylcysteine and sulforaphane, which act to increase glutathione, delayed the onset of epilepsy measured at 5 months without modifying the average seizure duration or the incidence of epilepsy in animals (
22). Considering the effect of vitamin E and the significant effect of auraptene on GSH at the highest dose, with anticonvulsant effect at lower doses, it may be suggested that the anti-epileptic effect of auraptene was mediated by a mechanism(s) other than modulation of the oxidant-antioxidant system.
It means that longer treatment with antioxidants may induce a better anticonvulsant effect. All these findings show that a combination of antioxidants with current anticonvulsant drugs is an alternative way that would be beneficial for the treatment of epilepsy.
Previous studies have suggested that seizures originate from two primary brain regions: the forebrain and the brainstem. These studies suggest that seizures characterized by forelimb clonus originate from forebrain structures such as the deep prepiriform cortex or the area tempestas, whereas tonic-clonic seizures are thought to originate from brainstem structures that include the pontine reticular formation and the nucleus reticularis pontis oralis (
23,
24). Therefore, it is a possibility that auraptene at the dose of 12.5 mg/kg had more inhibitory effect on the forebrain structures than higher doses and enhanced S2L more than other doses. Due to limited number of studies, the pharmacological effects of auraptene have not been evaluated in detail. Hence, it is hard to suggest the molecular mechanisms behind the anticonvulsant effect of auraptene. Similar to auraptene, there are other coumarin molecules such as esculetin (
25) and imperatorin (
26) with good anticonvulsant effects. Esculetin has been reported with significant anticonvulsant effect in electroshock-induced convulsions model without sedative and myorelaxant effects. Modulation of voltage-gated and ligand-gated ion channels is a potential mechanism for the anticonvulsant effects of plant compounds. It has been reported that coumarins interact with various voltage-gated ion channels and the benzodiazepine site of the GABAA receptors (
27,
28). In accordance, it was reported that the anticonvulsant effect of esculetin was mediated by GABAA receptors. The interaction of coumarin derivatives with GABAA receptors was evaluated by Singhuber
et al. (
29). They showed that these compounds enhance GABA-induced chloride current however, at very high concentrations. Therefore, it is unlikely that this target is the only mechanism that mediates the anticonvulsant effects of coumarins. Auraptene can reduce glutamate concentration as an excitatory neurotransmitter in the CNS (
5). This may be another mechanism that explains the anticonvulsant effect of auraptene in the present study. Auraptene has been reported as a ligand for PPARγ as well (
30). This nuclear receptor has various physiological effects including modulation of metabolism (
31) and inflammation (
32). Recent studies show that it is involved in the control of seizure episodes as well. For example, it was demonstrated that single intraperitoneal administration of pioglitazone, as a PPARγ agonist, reduced the proconvulsant effect of PTZ (
33,
34). The researchers suggested that PPARγ, through attenuation of proinflammatory cytokines and elevation of nitric oxide synthesis, induced anticonvulsant effects. Therefore, it is a possibility that auraptene induced its anticonvulsant effect through activation of PPARγ. There are reports showing that inflammation has a crucial role in the pathophysiology of epilepsy (
35). In agreement, it was demonstrated that the expression of cyclooxygenase-2 (COX-2) was elevated in the pyramidal cells of the hippocampus following kindling. Moreover, following kindling, prostaglandin E2 concentration did not increase in COX-2 knock-out mice despite a significant increase in the wild-type (
36). This implies the importance of COX-2 expression during epileptogenesis. Interestingly, there are reports showing that auraptene can change COX-2 expression. For example, auraptene inhibited COX-2 expression in the astrocytes and attenuated microglia activation in the hippocampus (
37). Similarly, Okuyama and colleagues showed that auraptene reduced COX-2, interleukin 1β, and TNF-α expressions in the ischemic brain of mice (
4). Therefore, another explanation for the anticonvulsant effect of auraptene is decreased expression of COX-2 and other pro-inflammatory chemokines and cytokines. It is worth to mention that IL-1β has a prominent place in the initiation and progression of PTZ-induced epileptogenesis (
38,
39).
Compounds with additional neuroprotective, cognition-enhancing, and antiinflammatory activities may be useful in the treatment of epilepsy. In accordance, Epifano
et al. demonstrated that auraptene produced a significant neuroprotective effect against N-methyl-D-aspartate (NMDA)-induced toxicity in mixed cortical cultures (
40). Auraptene has been reported with significant inhibitory action on MAO-B as a neuroprotective mechanism (
41). Moreover, auraptene improved reduced ischemia and enhanced memory in a rat model of vascular dementia (
14). On the other hand, one of the major pitfalls in the treatment of epilepsy is the interaction of the current antiepileptic drugs with hepatic drug-metabolizing enzymes and therefore with many other drugs. Interestingly, auraptene has been reported as a compound with the lowest interaction with these enzymes (
42). Therefore, the neuroprotective properties of auraptene and minimum interaction with hepatic drug-metabolizing enzymes may add benefits to the anticonvulsant effects of this almost safe (
43) natural compound.
In conclusion, auraptene induced significant anticonvulsant effect and increased reduced glutathione. Considering the weak anticonvulsant effect of vitamin E, it was suggested that mechanisms other than the antioxidant effect of auraptene participated in its anticonvulsant effects.