One of the most important results of our study was the role of TGFBR1 in ATLL. The TGFBR1 gene commands the synthesis of TGF-β receptor type 1 protein. The function of this receptor is to transmit signals from the cell surface into the cell via the signal transduction process (
13,
14). Since TGFBR1 prevents cells from growing and dividing too rapidly, it can function as a tumor suppressor (
15). We found that TGFBR1 gene expression is significantly higher in ATLL patients than in ACs and healthy donors. However, contrary to our results, some studies have demonstrated that decreased TGFBR1 signaling may contribute to cancer progression (
16,
17). We hypothesize that TGFBR1 expression increases to defend against cancer cells but cannot overcome them, resulting in higher gene expression than in healthy donors. Another possible explanation is the difference in the type of cancer.
The TGFBR2 gene encodes TGF-β receptor type 2, which is part of the serine/threonine protein kinase family. TGFBR2 is a transmembrane protein receptor with a protein kinase domain that forms a heterodimeric complex with another receptor protein and links TGF-beta. This receptor/ligand complex regulates cell proliferation through protein phosphorylation (
18,
19). Mutations in this gene are associated with various diseases, including the development of different types of tumors (
20). We found that the expression level of TGFBR2 in ATLL patients and ACs was significantly lower than in healthy donors. Our results suggest that TGFBR2 acts as a tumor suppressor. Consistent with our study, several studies have shown that a decrease in TGFBR2 expression level is associated with higher grades of cancer (
21-
23). We can conclude that in the early stages of cancer, TGFBR2 acts as a tumor suppressor.
One of the vital signaling networks is the transforming growth factor TGF‐β signaling pathway, which plays an essential role in cell proliferation, inflammation, apoptosis, angiogenesis, and tumor genesis (
24). Although the level of TGF-β is nearly the same in ATLL patients and healthy humans (
25),
in vitro studies show that in ATLL patients, the expression of TGF-β mRNA and secretion of TGF-β is higher than in healthy humans (
26-
28). While TGF-β usually inhibits cell proliferation (
29), it has no inhibitory effect on the proliferation of T-cells infected with HTLV-1 (
30).
Transforming growth factor-beta (TGFβ) is a pleiotropic multifunctional cytokine secreted from epithelial cells and fibroblasts (
31,
32). When TGF-β family ligands bind to the active TGFβ type II serine/threonine kinase receptor (TβRII), they recruit the type I receptor (TβRI) and form a stable oligomeric receptor complex. The formation of this complex causes the type II receptor to phosphorylate the type I receptor. This phosphorylation activates the type I receptor-kinase through structural changes. SMAD2/3 phosphorylation results in binding to SMAD4 and the creation of a signaling complex, which translocates to the nucleus to induce downstream changes in gene expression through the canonical signaling pathway.
After nuclear translocation, to regulate gene expression, the SMAD complex binds to other transcription factors or chromatin-remodeling proteins. Transforming growth factor-beta 1 also functions through a plethora of noncanonical SMAD-independent pathways, including the mitogen-activated protein (MAP) kinase pathway, which signals via extracellular signal–regulated kinase 1/2 or TGF-β–activated kinase 1 (TAK1) (
33,
34). Several studies have demonstrated that TGF-β is deregulated in cancers and plays an essential role in tumor initiation and progression. In the early stage of tumors, TGF-β acts as a tumor suppressor through its antiproliferative effects, while in the later stages, it acts as a tumor promoter, aiding tumor metastasis through an autocrine TGF-β loop (
35,
36).
Among the three TGF-β isoforms, TGF-β1 is the most essential, controlling differentiation, hemostasis, and the development of other immune cells. Transforming growth factor-beta 1 is a potent inhibitor of cell proliferation in most endothelial, epithelial, and hematopoietic cells, such as T lymphocytes. Transforming growth factor-beta 1 suppresses tumor cells in their early stages. However, tumor cells develop mechanisms to escape the inhibitory effects of TGF-β1. Some mutations occur in TGF-β receptors, for example, downstream in TGFBR1 and TGFBR2, SMAD2, and SMAD4, which occur in metastatic colon cancer and pancreatic cancer (
37).
Kim et al. found that in ATL cells, TGF-β is overproduced, and the main isoform secreted by these cells is TGF-β1. Cytokines such as IL-10 and TGF-β suppress the immune system in ATLL patients. Although the TGF-β level in the serum of ATLL patients is not significantly higher than in healthy people,
in vitro studies of ATLL cell culture and long-term ATLL T-cell lines have proven that ATLL cells express TGF-β mRNA and secrete TGF-β cytokines. Although TGF-β is a well-known cell proliferation inhibitor, it cannot prevent the spontaneous proliferation of HTLV-1-infected T cells. Kim et al. found that the expression of TGF-β1 in leukemic cells of ATLL patients and HTLV-1-infected T-cells is high (
28).
Following our study, Javle et al. found that the TGF-β signaling pathway plays an important role in pancreatic cancer. They demonstrated that the TGF-β1 plasma level was high in pancreatic cancer; however, the expression level of TGFBR2 and SMAD4 was not related to pancreatic cancer (
38). Hadi et al. showed that TGFBR1 and TGFBR2 have crucial roles in chronic lymphocytic leukemia (CLL), and five deregulated miRNAs, including mir-574, mir-499, mir-125b, mir-106a, and mir-9, are involved in CLL pathogenesis (
39).
Mir-20a-5p is a component of the mir-17-92 cluster, which is associated with many kinds of cancers. Overexpression or downregulation of miR-20a-5p participates in downstream signaling pathways, such as PI3K-Akt, MAPK, and TGF-β signaling pathways. Controlling incessant proliferation signals, evading growth inhibition, activating invasion and metastasis, enabling limitless replication, promoting angiogenesis, resisting cellular death, and avoiding immune destruction are mechanisms of miR-20a-5p (
12). Wei Huang et al. have demonstrated that deregulated miRNAs are one of the vital causes of cancers, making them biomarkers for cancer diagnosis and prognosis (
40).
In this study, we identified the TGF-β signaling pathway as the most important signaling pathway in ATLL pathogenesis. Mohadeseh Valizadeh et al. demonstrated that downregulation in the TGF-β signaling pathway is associated with sulfur mustard (SM) skin lesions. They showed that TGF-β, TGFBR2, and their functional miRNAs, including mir-20a and mir-21a, contribute to SM lesions (
41). Zhang et al. determined that mir-20a is an independent prognostic and therapeutic factor for colorectal cancer (
42). In this study, we also demonstrated that SMAD4, BMPR2, RBL1, ZFYVE9, BAMBI, MAPK1, RGMB, E2F5, SMAD5, RGMA, and TGFBR2 genes are involved in the TGF-β signaling pathway. TGF-beta family members, which include TGF-betas, bone morphogenetic proteins (BMPs), and activins, are structurally related secreted cytokines with a broad spectrum of cellular functions (
43,
44).
5.1. Conclusions
In this study, which resulted from the bioinformatic analysis of high-throughput microarray data, it was determined that TGFBR1 has a more active role than TGFBR2 and even TGF-beta itself. Additionally, Mir-20a-5p and the genes SMAD4, BMPR2, RBL1, ZFYVE9, BAMBI, MAPK1, RGMB, E2F5, SMAD5, RGMA, and TGFBR2 were identified as main genes in the TGF-beta signaling pathway. More studies are needed to further clarify the role of TGFBR1.
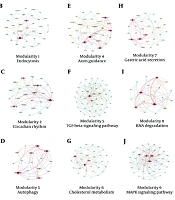







 in human T-cell lymphotropic virus type 1 (HTLV-1) infected cases; B, modularity 1: Endocytosis; C, modularity 2: Circadian rhythm pathway; D, modularity 3: Autophagy pathway; E, modularity 4: Axon guidance pathway; F, modularity 5: Transforming growth factor-beta (TGF-β) pathway; G, modularity 6: Cholesterol metabolism; H, modularity 7: Gastric acid secretion pathway; I, modularity 8: RNA degeneration pathway; J, modularity 9: MAPK signaling pathway.")
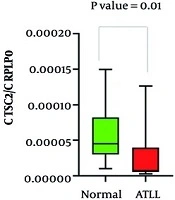
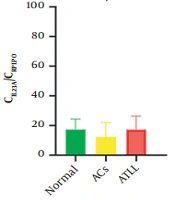
 among healthy, asymptomatic carriers (ACs) and adult T-cell leukemia/lymphoma (ATLL); B, expression level of TGFBR1 among healthy, ACs and ATLL; C, expression level of TGFBR2 among healthy, ACs and ATLL")