In this study, the effects of MLR-1023 on cognitive deficiency and oxidative stress in an LPS-induced mouse model of AD were examined. To investigate the possible involvement of the NO/cGMP/KATP channels pathway and PPARγ receptors in the effects of MLR-1023, mice were pre-treated with related agonists and antagonists that did not have a significant effect when used alone.
It has been shown that LPS induces inflammation that can lead to memory impairment and hippocampal-related disabilities (
22). Our results indicated that LPS injection increased the initial IL, reflecting cognitive impairment, and decreased the STL time, reflecting learning-memory impairment, in the passive avoidance task in the shuttle box test. MLR-1023 decreased IL and increased STL at the dose of 40 mg/kg. Thus, we can conclude that MLR-1023 can improve cognitive deficiency and learning-memory impairment induced by LPS.
NO, as a key neurotransmitter, is involved in neurodegenerative disorders such as cognitive impairments (
23). It has been reported that the NO-cGMP pathway is involved in some spatial and recognition memory impairments (
24). In this study, pre-treatment with L-NAME increased IL and decreased STL, indicating that L-NAME reduced the improving effect of MLR-1023. In contrast, pre-treatment with L-arginine decreased IL and increased STL, showing that L-arginine potentiated the improving effect of MLR-1023. Therefore, it can be inferred that the L-arginine-NO pathway may be involved in the improving effect of MLR-1023 on cognitive deficiency and learning-memory impairments induced by LPS in the passive avoidance task in the shuttle box test.
Previous studies confirm that the NO/cGMP signaling pathway is involved in the learning-memory process. NO plays a crucial role in long-term potentiation and synaptic plasticity, which are essential for learning and memory activities. These evidences suggest that NO could provide a therapeutic opportunity in AD via modulation of the relevant NOS expression (
23). In this regard, we observed that pre-treatment with methylene blue increased IL and decreased STL, indicating that methylene blue reduced the improving effect of MLR-1023. In contrast, sildenafil decreased IL and increased STL, enhancing the improving effect of MLR-1023. Thus, guanylate cyclase (GC) and cGMP may play a role in the improving effect of MLR-1023 on cognitive deficiency and learning-memory impairments induced by LPS in the shuttle box test.
K
ATP channels regulate many cellular functions by connecting metabolic function and cell membrane electrical functions. Opening K
ATP channels can decrease brain damage, improve learning and memory, and provide a neuroprotective role by stimulating synaptic connections (
25). In this study, pretreatment with glibenclamide (a K
ATP channel blocker) increased the IL and decreased the STL, indicating that it weakens the improving effect of MLR-1023. In contrast, diazoxide (a K
ATP channel opener) decreased the IL and increased the STL, enhancing the improving effect of MLR-1023. Thus, K
ATP channels may play a role in the improving effect of MLR-1023 on cognitive deficiency and learning-memory impairments induced by LPS in the shuttle box test.
Studies have shown that dysfunction of PPAR-γ receptors during neurodevelopment can cause memory impairment, whereas PPAR-γ activation is a potential therapeutic approach for these problems (
26). However, our results showed that pretreatment with pioglitazone (a PPAR
γ receptor agonist) and GW9662 (a PPAR
γ receptor antagonist) did not have any significant effect on the passive avoidance task in the shuttle box test. Thus, PPAR
γ receptors did not play a role in the improving effect of MLR-1023 on cognitive deficiency and learning-memory impairments induced by LPS. These agonists and antagonists did not significantly affect IL and STL time when used alone.
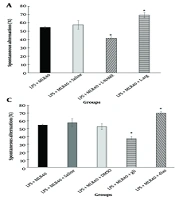
Lipopolysaccharide can cause reactive oxygen species (ROS)-mediated rat hippocampal neuroinflammation, leading to memory loss (
27). In our study, LPS injection significantly decreased the spontaneous alternation percentage compared to the control group in the Y-maze test, indicating learning-memory impairment. Treatment with MLR-1023 increased the spontaneous alternation percentage, showing the improving effect of MLR-1023 on learning-memory impairments induced by LPS. There was no difference in total arm entries between groups, indicating that the mice did not have motion impairment.
Several studies reveal that activation of neuronal nitric oxide synthase leads to increased NO levels, which activate guanylate cyclase, in turn increasing cGMP levels. A deficiency of this NO-cGMP pathway reduces the ability to learn a Y-maze task. Thus, the NO-cGMP pathway modulates the ability of rats to learn the Y-maze task, and cognitive ability in the Y-maze test may be improved by enhancing this pathway (
28). Similarly, our results showed that pretreatment with L-NAME decreased the spontaneous alternation percentage compared to the LPS + MLR-1023 treated group, decreasing the improving effect of MLR-1023 on learning-memory impairments induced by LPS. In contrast, L-arginine increased the spontaneous alternation percentage, showing it enhanced the improving effect of MLR-1023 in the Y-maze test. Therefore, we can infer that the L-arginine-NO pathway may be involved in the improving effect of MLR-1023 on learning-memory impairments induced by LPS. Furthermore, pretreatment with methylene blue decreased the spontaneous alternation percentage, decreasing the improving effect of MLR-1023 on learning-memory impairments induced by LPS. On the other hand, sildenafil increased the spontaneous alternation percentage, potentiating the improving effect of MLR-1023 in the Y-maze test. Thus, GC and cGMP may be involved in the improving effect of MLR-1023 on learning-memory impairments induced by LPS. Supporting this, previous studies have proposed that extracellular cGMP modulates the ability to learn Y-maze behavior (
29).
We observed that pretreatment with glibenclamide (a K
ATP channel blocker) decreased the spontaneous alternation percentage, thus diminishing the improving effect of MLR-1023 on learning-memory impairments induced by LPS. Similarly, some previous findings indicate that glibenclamide enhances spontaneous alternation performance, suggesting that ATP-sensitive potassium channels may play a role in cognitive performance (
30). Conversely, pretreatment with diazoxide (a K
ATP channel opener) increased the spontaneous alternation percentage, enhancing the improving effect of MLR-1023 in the Y-maze test. Overall, it appears that K
ATP channels may be involved in the improving effect of MLR-1023 on learning-memory impairments induced by LPS.
It has been reported that PPAR
γ receptors play an important role in the Y-maze task analysis. As pioglitazone, a PPAR
γ agonist, can improve type 2 diabetes-related cognitive insufficiencies, it can be concluded that pioglitazone may be considered a potential agent for managing cognitive deficiencies (
31). In our study, pioglitazone increased the spontaneous alternation percentage, thus enhancing the improving effect of MLR-1023 in the Y-maze test. However, pretreatment with GW9662 (a PPAR
γ receptor antagonist) decreased the spontaneous alternation percentage, indicating a reduction in the improving effect of MLR-1023 on learning-memory impairments induced by LPS. Therefore, PPAR
γ receptors may play a role in the improving effect of MLR-1023 on learning-memory impairments induced by LPS. These agonists and antagonists did not have a significant effect on the spontaneous alternation percentage when used alone.
A study has shown that antioxidant capacity, such as SOD activity, decreased in the brain of LPS-treated rats, while lipid peroxidation (MDA) and cytokine production increased (
32). In our study, LPS injection significantly increased the levels of MDA, a lipid peroxidation marker, in hippocampal tissue. Treatment with MLR-1023 decreased the levels of MDA, demonstrating an improving effect on lipid peroxidation induced by LPS in hippocampal tissue.
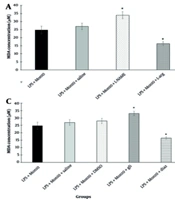
Our study showed that pretreatment with L-NAME increased the levels of MDA, reducing the improving effect of MLR-1023 on lipid peroxidation induced by LPS. Conversely, pretreatment with L-arginine decreased the levels of MDA, enhancing the improving effect of MLR-1023 on lipid peroxidation induced by LPS. Thus, we infer that the L-arginine-NO pathway may have a possible involvement in the improving effect of MLR-1023 on lipid peroxidation induced by LPS in hippocampal tissue. Additionally, pretreatment with methylene blue increased the levels of MDA, indicating a reduction in the improving effect of MLR-1023 on LPS-induced lipid peroxidation. However, pretreatment with sildenafil did not have a significant effect on the levels of MDA. Nevertheless, GC and cGMP may have a possible role in the improving effect of MLR-1023 on LPS-induced lipid peroxidation in hippocampal tissue. One of the notable pathological changes in AD patients is lipid peroxidation. Furthermore, malondialdehyde, as an end product of peroxidation, is also increased in the brain of patients with AD (
33).
Studies have shown that K
ATP channel blockage by glibenclamide reduces the beneficial effects of simvastatin on MDA content. Therefore, the opening of K
ATP channels plays a crucial role in protection against lipid peroxidation (
34). In this project, pretreatment with glibenclamide increased MDA levels, diminishing the improving effect of MLR-1023 on LPS-induced lipid peroxidation. In contrast, pretreatment with diazoxide (a K
ATP channel opener) decreased MDA levels, enhancing the effect of MLR-1023. Thus, K
ATP channels may have a potential role in the positive effect of MLR-1023 on LPS-induced lipid peroxidation in hippocampal tissue.
Our results indicate that pretreatment with pioglitazone (a PPAR
γ receptor agonist) decreased MDA levels, enhancing the improving effect of MLR-1023 on LPS-induced lipid peroxidation. However, pretreatment with GW9662 (a PPAR
γ receptor antagonist) did not significantly affect MDA levels. Therefore, PPAR
γ receptors may have a potential role in the beneficial effect of MLR-1023 on LPS-induced lipid peroxidation in hippocampal tissue. These agonists and antagonists did not significantly affect MDA levels when used alone. However, another investigation shows that PPAR
γ receptors can induce the accumulation of lipid droplets, indicating the molecular regulatory mechanism of PPAR
γ on lipid metabolism (
35).
Results from a study revealed that LPS increased MDA levels and decreased SOD activity. IL-1β knockdown in the hippocampus improved oxidative stress and neuroinflammation induced by LPS, reducing memory insufficiencies and depression-like behaviors. This suggests that IL-1β is necessary for the oxidative and neuroinflammatory responses produced by LPS (
36). In our study, LPS decreased SOD enzyme activity (u/mL) in hippocampal tissue, indicating a reduction in antioxidative capacity and oxidative stress. Treatment with MLR-1023 increased SOD activity, demonstrating that MLR-1023 can improve LPS-induced impairment in SOD activity in hippocampal tissue.
Our results revealed that pretreatment with L-NAME decreased SOD activity, reducing the beneficial effect of MLR-1023 on LPS-induced impairment in SOD activity. In contrast, pretreatment with L-arginine did not significantly affect SOD activity, suggesting that the L-arginine-NO pathway may play a potential role in the improving effect of MLR-1023 on LPS-induced impairment in the antioxidative capacity of hippocampal tissue. Moreover, pretreatment with methylene blue decreased SOD activity, reducing the improving effect of MLR-1023 on LPS-induced impairment in SOD activity. However, pretreatment with sildenafil increased SOD activity, enhancing the effect of MLR-1023 on LPS-induced impairment in SOD activity. Thus, GC and cGMP may be involved in the improving effect of MLR-1023 on LPS-induced impairment in the hippocampal tissue antioxidative capacity.
Pretreatment with glibenclamide (a KATP channel blocker) decreased SOD activity, reducing the improving effect of MLR-1023 on LPS-induced impairment in SOD activity. Conversely, pretreatment with diazoxide (a KATP channel opener) did not significantly affect hippocampal tissue SOD activity, suggesting that KATP channels may play a potential role in the improving effect of MLR-1023 on LPS-induced impairment in hippocampal tissue antioxidative capacity.
Pretreatment with pioglitazone (a PPARγ receptor agonist) increased SOD activity, enhancing the effect of MLR-1023 on LPS-induced impairment in SOD activity. However, pretreatment with GW9662 (a PPARγ receptor antagonist) decreased SOD activity, reducing the improving effect of MLR-1023 on LPS-induced impairment in SOD activity. Consequently, PPARγ receptors may play a role in the beneficial effect of MLR-1023 on LPS-induced impairment in hippocampal tissue antioxidative capacity. These agonists and antagonists did not significantly affect hippocampal tissue SOD activity when used alone.
In conclusion, MLR-1023 can improve learning-memory impairments and oxidative stress induced by LPS in a mice model. The KATP/cGMP/NO cell signaling pathway and PPARγ receptors may be involved in this effect.






 in the initial latency (IL) and step-through latency (STL) time in the shuttle box test (A); spontaneous alternation percentage in Y-maze test (B) versus control group in mice. Values are expressed as mean ± SEM. (n = 6). * P < 0.05 and *** P < 0.001 compared with the control group (one-way ANOVA followed by Tukey's test). # P < 0.05 and ## P < 0.01 compared with the lipopolysaccharide (LPS) + vehicle treated group (one-way ANOVA followed by Tukey's test).")
/K<sub>ATP</sub> channels pathway and PPAR<sub>γ</sub> receptors in the effect of MLR-1023 on the initial latency (IL) and step-through latency (STL) in shuttle box test, are shown in panels A, B, C and D respectively. Values are expressed as mean ± SEM. (n = 6). * P < 0.05, ** P < 0.01 compared with the lipopolysaccharide (LPS) + MLR-1023 (40 mg/kg, i.p.) treated group, as determined by one-way ANOVA followed by Tukey’s (LSD in some cases) post hoc test.")
/K<sub>ATP</sub> channels pathway and PPAR<sub>γ</sub> receptors in the effect of MLR-1023 on learning-memory in Y-maze test, are shown in panels A, B, C and D respectively. Values are expressed as mean ± SEM. (n = 6). * P < 0.05, ** P < 0.01 compared with the lipopolysaccharide (LPS) + MLR-1023 (40 mg/kg, i.p.) treated group, as determined by one-way ANOVA followed by Tukey’s post hoc test.")
 on the levels of malondialdehyde (MDA) (A); and superoxide dismutase (SOD) enzyme activity (B) versus control group in the mice brain tissue. Values are expressed as mean ± SEM. (n = 6). *** P < 0.001 compared with the vehicle-treated control group (one-way ANOVA followed by Tukey's test). # P < 0.05, ## P < 0.01 compared with the lipopolysaccharide (LPS) + vehicle treated group (one-way ANOVA followed by Tukey's test).")
![Involvement NO/cyclic GMP (cGMP)/K<sub>ATP</sub> channels pathway and PPAR<sub>γ</sub> receptors in the effect of MLR-1023 on hippocampal tissue lipid peroxidation [levels of malondialdehyde (MDA)] are shown in panels A, B, C and D respectively. Values are expressed as mean ± SEM. (n = 6). * P < 0.05, ** P < 0.01 compared with the lipopolysaccharide (LPS) + MLR-1023 (40 mg/kg, i.p.) treated group, as determined by one-way ANOVA followed by Tukey’s post hoc test.](https://brieflands.com/journals/jai/articles/149170/figures/jai-149170-i006-F6-preview.webp "Involvement NO/cyclic GMP (cGMP)/K<sub>ATP</sub> channels pathway and PPAR<sub>γ</sub> receptors in the effect of MLR-1023 on hippocampal tissue lipid peroxidation [levels of malondialdehyde (MDA)] are shown in panels A, B, C and D respectively. Values are expressed as mean ± SEM. (n = 6). * P < 0.05, ** P < 0.01 compared with the lipopolysaccharide (LPS) + MLR-1023 (40 mg/kg, i.p.) treated group, as determined by one-way ANOVA followed by Tukey’s post hoc test.")
/K<sub>ATP</sub> channels pathway and PPAR<sub>γ</sub> receptors in the effect of MLR-1023 on hippocampal tissue superoxide dismutase (SOD) activity are shown in panels A, B, C and D respectively. Values are expressed as mean ± SEM. (n = 6). * P < 0.05 compared with the lipopolysaccharide (LPS) + MLR-1023 (40 mg/kg, i.p.) treated group, as determined by one-way ANOVA followed by Tukey’s post hoc test.")