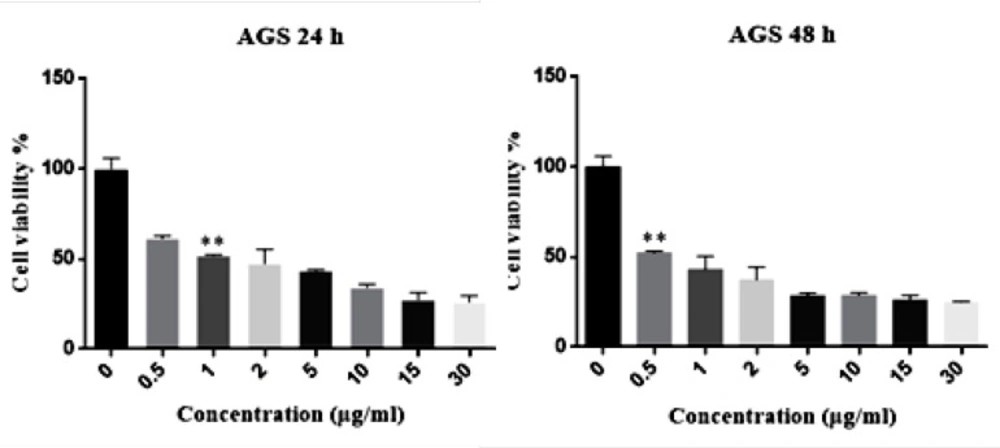
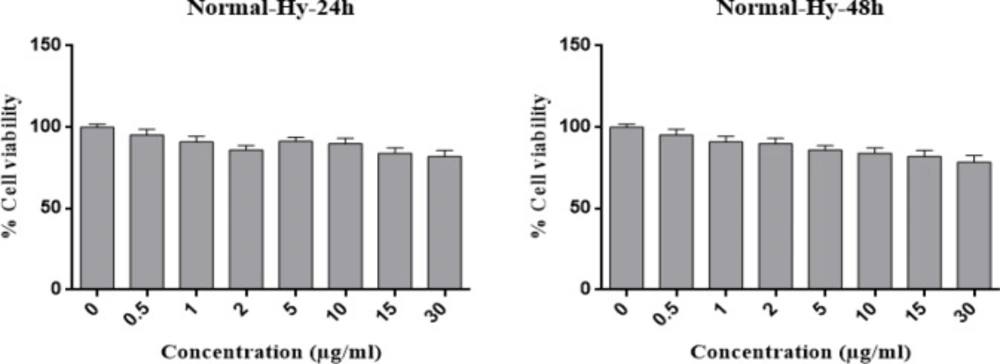
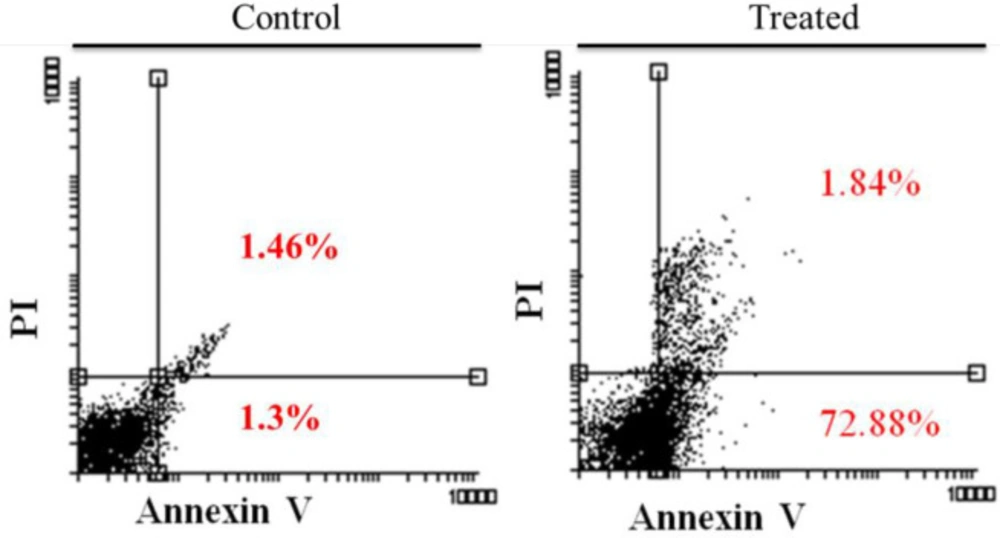
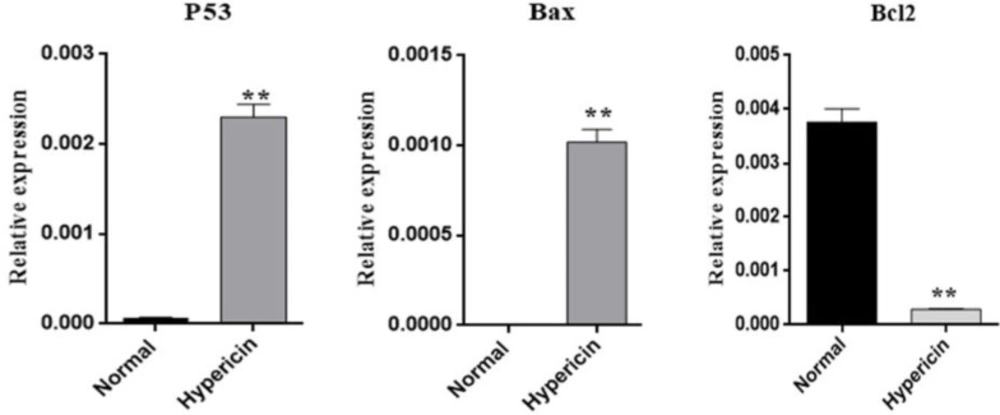
Gastric cancer is a common cancer with high mortality rate (
17). The treatment of gastric cancer is done by surgery, radiotherapy, and chemotherapy. However, tumor resistance to radiotherapy and chemotherapy calls for development of novel anticancer drugs (
9,
10,
18).
Hypericum perforatum is a medicinal plant, having hypericin as one of its active ingredients. Previous studies have shown that hypericin has antioxidant effects and is cytotoxic to some cancer cell lines (
17,
19).
In a study by Mirmalek
et al. on the effect of Hypericin on breast cancer cell line, they concluded that hypericin played a dose-dependent apoptotic impact on MCF-7 cell line. They showed that the IC50 hypericin and cisplatin on the MCF-7 cell line were 5 μg/mL and 20 μg/mL, respectively, indicating the effectiveness of hypericin at much lower concentrations compared with cisplatin. Moreover, they concluded that hypericin had a suitable cytotoxic effect on the breast cancer cell line and is a good candidate to be used in the treatment of breast cancer (
17). Hamilton
et al. showed that hypericin induced apoptosis in hypophysis adenoma cells AtT-20 and GH4C1 at 100 nM. Interestingly, a concentration of 10 μM did not induce apoptosis in human fibroblasts (
20). Moreover, Kim
et al. (
21) demonstrated that that concentration of hypericin for growth inhibition in 50% of histiocytic lymphoma cells was 0.2 µM. Yi
et al. (
22) found that photoactivated hypericin at a concentration of 50 nM induced apoptosis through elevation of
Bax-to
Bcl-2 ratio in RINm5F insulinoma cells. In the study caspase-3 and caspase-9 were also found to be elevated. In addition to inducing apoptosis, hypericin has also been reported to eliminate von-Hippel Lindau protein from cancerous cells and thus prevented growth of cancer cells (
23). In contrast to this study and other reports on the apoptotic cell death caused by hypericin, Mikeš
et al. (
24) found that necrosis is the main type of cell death in human colon adenocarcinoma HT-29 cells treated with photodynamic therapy with 60-400 nM hypericin. Another report in which did not confirm our study was done by Do
et al. (
25). They found that hypericin, at a concentration of 0.5 μM, inhibited methylglyoxal-induced apoptosis. The action was mediated through changes in the expression level of
Bcl-2 and
Bax.
We observed down-regulation of
Bcl-2 and up-regulation of Bax. Photoactivated hypericin (0.021 μM) in presence of genistein also down-regulated
Bcl-2 and up-regulated Bax in human breast adenocarcinoma cell lines MCF-7 and MDA-MB-231 (
26). In a study by You
et al. (
27) the extract of
Hypericom perforatum, the source of hypericin
, also increased the expression of Bax and decreased the expression of
Bcl-2.
Hypericin may physically interact with
Bcl2 family proteins (
28)). In U87 MG and HCAEC cells, hypericin (500 nM) significantly changed the distribution of
Bcl2 and
Bax proteins in the dark (
29). Hypericin also alters the distribution of
Bcl-2 in U-87 MG glioma cells (
30) and in human coronary aorta endothelial cells (
31). In the study by Chan
et al. (
32), six hour post hypericin photodynamic therapy, apoptotic nuclei were seen in HK-1 nasopharyngeal carcinoma cells.
Bax translocation and formation of
Bax channels in mitochondria membrane were mentioned as the causes of cell death in the study. As found by Balogová (
30) apoptotic stimulus by hypericin photodynamic action on U-87 MG glioma cell line caused Bax translocation into mitochondria. In sonodynamic therapy with hypericin (25 μg/mL) in THP-1 macrophages
Bcl-2 protein was found to be down-regulated (
33).
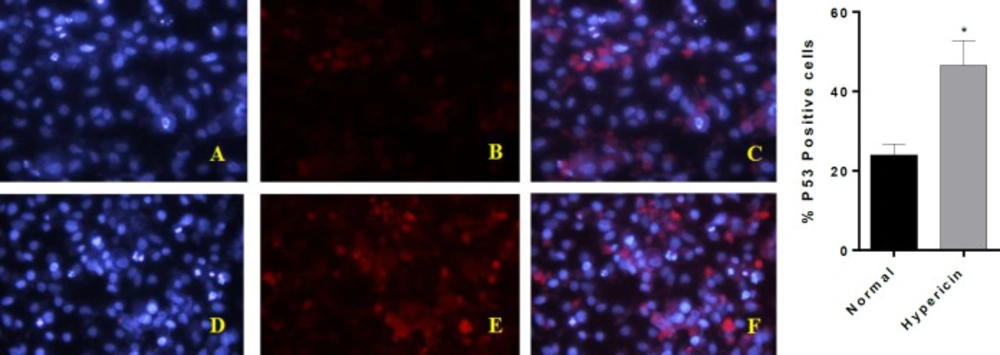
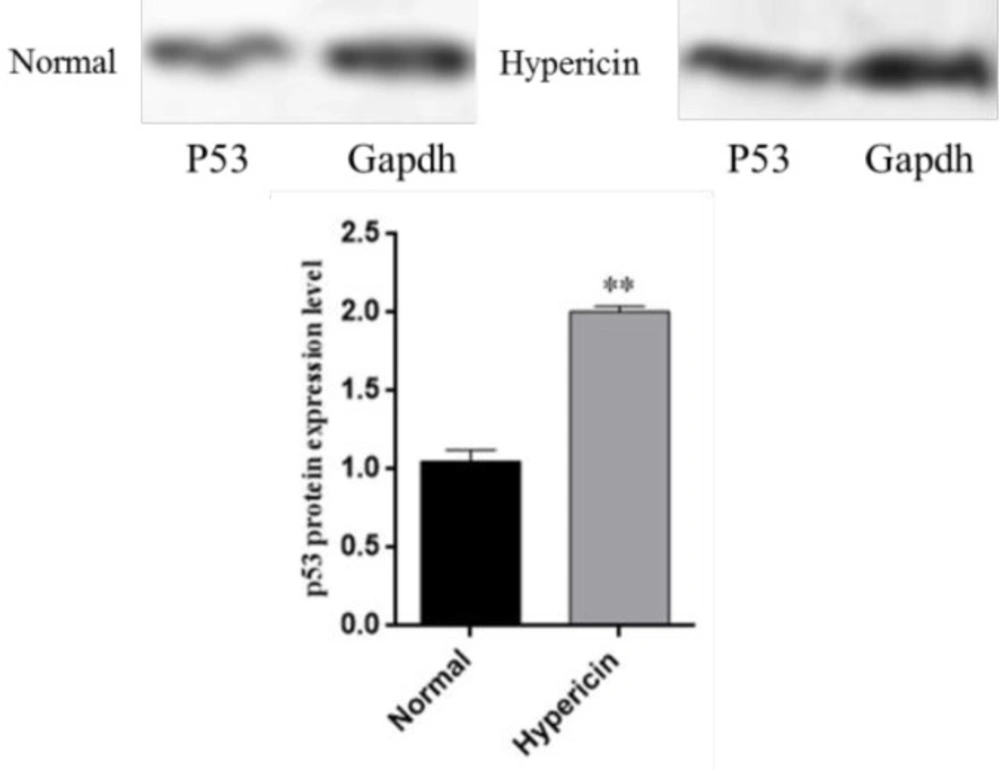
In a survey by Acar
et al. (
34) it was shown that treatment of MCF-7 breast cancer cells with hypericin (7.5, 5 and 1 μM ) did not alter expression of
p53 gene, although it did induce apoptosis. Furthermore, the level of
Bcl-2 was not altered in both
p53-null and wt-
p53-expressing HCT-116 cells after hypericin photodynamic therapy, which is in contrast to our findings (
35).
Xu
et al.(
36) found that hypericin (6.25-200 ng/mL) photodynamic therapy induced apoptosis and G2/M phase cell cycle arrest in Adult T-cell leukemia.
Bcl-2 was down-regulated while Bax and p53 were up-regulated. Gamasaee
et al. (
37) found that
p53 was up-regulated in MDA-MB-175-VII cells which underwent apoptosis when treated with hypericin. Halaburková
et al.(
38) showed that the induction of apoptosis in colon cancer cells HT-29 and HCT 116 co-treated with the histone deacetylase inhibitor sodium phenylbutyrate+ hypericine was
P53-dependent. The authors also suggested that the effect was due to hypericin photodynamic therapy rather than sodium phenylbutyrate. These results are also in line with our findings. However, the results by Lee
et al. suggested that hypericin (0.06-1 µ/mL) mediated phototherapy acted independently from p53, and the authors suggested that photodynamic therapy may have equal efficacy regardless of the presence or absence of
p53 (
39). In a similar report Weller
et al. (
40) had found that in human malignant glioma cells, hypericin induced apoptosis independently of
p53.
Finally we suggest that the synergistic effect of YM155 (
29) or genistein (
26) with hypericin may be also tested in induction of apoptosis in AGS cells.